
RNA sequencing (RNA-Seq)
RNA-SEQ: EXPLORING BEYOND THE COMMONLY STUDIED MODEL ORGANISMS
RNA sequencing (RNA-Seq) has revolutionized the study of gene expression and regulation, thus providing deep insights into the inner workings of living organisms. One common method for this technology, total RNA-Seq, profiles both coding and noncoding RNA with the depletion of the overly abundant ribosomal RNA (rRNA). With most existing rRNA depletion methods customized and established for human samples and the commonly studied mouse and rat model systems, the Zymo-Seq RiboFree® Total RNA Library Kit allows researchers to expand their horizons thanks to its novel rRNA depletion technology that is probe-free and species-independent. In this blog, you will find several recently published studies where researchers have leveraged the RiboFree® depletion strategy to explore the transcriptomes of organisms outside the common models.
THE STUDY OF THE CRIMEAN-CONGO HEMORRHAGIC FEVER VIRUS (CCHFV)

As the scientific community transitions into a post-COVID era, there is an increased focus on epidemiological studies involving potential sources of zoonotic viruses. A recent study conducted by scientists in Uganda shed light on an insufficiently studied yet widespread tickborne pathogen called the Crimean-Congo hemorrhagic fever virus (CCHFV). Amidst their investigation, the researchers employed the Zymo-Seq RiboFree® Total RNA Library Kit to analyze a novel viral strain of the CCHFV.1
CCHFV is transmitted between livestock and humans through infected tick bites, causing severe outbreaks across Africa, Asia, the Middle East, and Eastern Europe.2,3 Despite being the most geographically widespread tick-borne viruses, strains of CCHFV remained poorly studied, especially true for the African strains, with most knowledge derived solely from severe human disease cases.
In this study, researchers took a novel approach by collecting RNA samples directly from the infected African blue ticks, Rhipicephalus (Boophilus) decoloratus. The aim was to expand the understanding of the natural variation of CCHFV strains from tick vectors and animal reservoirs directly. The Zymo-Seq RiboFree® Total RNA Library Kit enabled the authors to successfully generate total RNA libraries from tick RNA for sequencing. From there they were able to characterize the complete coding region of this novel CCHFV strain, contributing to the growing reservoir of data essential for the development of vaccines, diagnostic tools, and control strategies for combating viral infections.1
THE STUDY OF THE MUSTARD HILL CORAL, PORITES ASTREOIDES

Coral reefs harbor 25% of the world’s biodiversity and unfortunately face significant challenges due to climate change-induced thermal stress.4, 5 This has led to a decline in global coral populations.5 While some reef species are highly sensitive to these thermal changes, others demonstrate remarkable resilience and even appear to thrive under these normally unfavorable conditions. One such resilient species is the mustard hill coral (Porites astreoides).6
To unravel the mechanisms behind the mustard hill coral’s resilience, researchers from the University of Rhode Island utilized the innovative Zymo-Seq RiboFree® technology to generate an ab initio reference transcriptome of P. astreoides, adding a useful resource for future studies.7
The researchers further leveraged the RNA-Seq data from the RiboFree® libraries to characterize the mapping potential of the draft reference genome they built from DNA sequencing. Remarkably, they achieved alignment rates of around or above 80% for unique sequences. This high alignment rate underscores the suitability of the new reference genome for future transcriptomic studies. As climate change continues to impact global biodiversity, understanding how resilient species like the mustard hill coral cope with these unprecedented challenges offers crucial insights into the future of our ecosystems.
THE STUDY OF OENOCOCCUS OENI

Wine production is ironically very far from soothing; in fact, it’s quite a complex process. The quality and stability of the wine is dependent upon bacteria that are utilized for various biochemical reactions. One critical process in winemaking is malolactic fermentation (MLF)8, which plays a vital role in reducing wine acidity and enhancing microbiological stability9, both of which are highly desirable characteristics. Oenococcus oeni, a gram-positive heterofermentative lactic acid bacterium (LAB) species, is commonly used for controlled MLF in wine production due to its acid tolerance of high ethanol levels.10 However, its sensitivity to sulfur dioxide (SO2), an antiseptic compound commonly used in winemaking9, remains poorly understood.
To shed light on the transcriptional response of O. oeni during MLF under the stress of SO2 exposure, researchers from Australia conducted a comprehensive investigation.11 They utilized the Zymo-Seq RiboFree® Total RNA Library Kit to prepare total RNA-Seq libraries from O. oeni under various experimental conditions. Through sequencing and differential gene expression analysis, they unveiled key transcriptional changes induced by SO2 exposure, highlighting its potential as a target for the development of SO2-tolerant strains. These advancements are pivotal in enhancing wine production and expanding the knowledge within the winemaking community.
CONCLUSION
The significance of an RNA-Seq kit that is compatible across all species cannot be overstated. With the Zymo-Seq RiboFree® Total RNA Library Kit, scientists worldwide now have a powerful tool at their disposal to delve into the transcriptomics of less commonly studied organisms. As exemplified by the peer-reviewed research presented above, the possibilities for remarkable and unprecedented discoveries are unlimited. From unraveling the secrets of resilient coral species to untangling the intricate responses of bacteria in wine production, the Zymo-Seq RiboFree® Total RNA Library Kit has served as a key that opens the door to a wealth of knowledge and innovation.
REFERENCES
- Wampande, E. M.; Waiswa, P.; Allen, D. J.; Hewson, R.; Frost, S. D. W.; Stubbs, S. C. B. Phylogenetic Characterization of Crimean-Congo Hemorrhagic Fever Virus Detected in African Blue Ticks Feeding on Cattle in a Ugandan Abattoir. Microorganisms 2021, 9 (2). DOI: 10.3390/microorganisms9020438.
- Bente, D. A.; Forrester, N. L.; Watts, D. M.; McAuley, A. J.; Whitehouse, C. A.; Bray, M. Crimean-Congo hemorrhagic fever: history, epidemiology, pathogenesis, clinical syndrome and genetic diversity. Antiviral Res 2013, 100 (1), 159-189. DOI: 10.1016/j.antiviral.2013.07.006.
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J Med Ento mol 1979, 15 (4), 307-417. DOI: 10.1093/jmedent/15.4.307.
- Hughes, T. P.; Kerry, J. T.; Connolly, S. R.; Álvarez-Romero, J. G.; Eakin, C. M.; Heron, S. F.; Gonzalez, M. A.; Moneghetti, J. Emergent properties in the responses of tropical corals to recurrent climate extremes. Curr Biol 2021, 31 (23), 5393-5399. e5393. DOI: 10.1016/j.cub.2021.10.046.
- Hughes, T. P.; Anderson, K. D.; Connolly, S. R.; Heron, S. F.; Kerry, J. T.; Lough, J. M.; Baird, A. H.; Baum, J. K.; Berumen, M. L.; Bridge, T. C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359 (6371), 80-83. DOI: 10.1126/science.aan8048.
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; van Woesik, R. Coral bleaching: the winners and the losers. Ecology Letters 2001, 4 (2), 122-131.
- Wong, K. H.; Putnam, H. M. The genome of the mustard hill coral, Porites astreoides. GigaByte 2022, 2022, gigabyte65. DOI: 10.46471/gigabyte.65
- Ribereau-Gayon, P.; Dubourdieu, D.; Doneche, B.; Lonvaud, A. Handbook of Enology: The Microbiology of Wine and Vinifi cations; 2006.
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie Van Leeuwenhoek 1999, 76 (1-4), 317-331.
- Henick-Kling, T. Malolactic Fermentation. In Wine Microbiology and Biotechnology, Fleet, G. H. Ed.; 1993; pp 289-326.
- Onetto, C. A.; Costello, P. J.; Kolouchova, R.; Jordans, C.; McCarthy, J.; Schmidt, S. A. Analysis of Transcriptomic Response to SO2 by Oenococcus oeni Growing in Continuous Culture. Microbiol Spectr 2021, 9 (2), e0115421. DOI: 10.1128/ Spectrum.01154-21.
- RNA-SEQ: EXPLORING BEYOND THE COMMONLY STUDIED MODEL ORGANISMS
October 11, 2023
Testimonials
Testimonials

During the 7 years that we have worked with Biolinkk as our regional distributor in India they have been well-planned and goal-oriented partners with Solis BioDyne, who always offer excellent customer service and ensure great customer satisfaction.

Carolin Kuuskmäe
Marketing Specialist
Biolinkk has been a representative of Molecular Research Center, Inc in India since 2008. We are very pleased with their performance and can recommend them as a competent partner in the global business world.
Shirley Pohlman
Molecular Research Center, Inc.

With the all-time fast response customer service and tech support, Biolinkk successfully makes their company stand out from a very competitive market. We believe Biolinkk can provide the best shopping experience for all our customers in India.
Wei "Amanda" Mao
Apex Biotechnology LLC

M/s.Biolinkk has a good nationwide network in India and provide best supports with researchers. We appreciate all supports of M/s. Biolinkk.

Masaki
Funakoshi Co. Ltd.

It is a pleasure to working with Biolinkk and must compliment on all of your efforts for developing the marketing. We would love to you feature us on your website. We will provide a better service and supports our good partner the best.
Ariel Chen
GeneDireX, Inc.)

Biolinkk is a reliable distributor. Very good cooperation with them
B.RGDS Scarlett
Miulab

We have worked with Biolinkk for over five years in the promotion of our Benchtop equipment. They have always provided a great deal of focus and knowledge related to our products as well as an exceptional level of support.
Tony Demsia
Benchmark Scientific Inc.

Professional, aggressive! Customer First! Always bear in mind the interest of the customers.
Tony
Blue-ray Biotech Corporation

Arab Lab Live 23



A Deep Dive into the Hidden Power of Programmed Cell Death: Unlocking Therapeutic Potential with Antibodies

A Deep Dive into the Hidden Power of Programmed Cell Death: Unlocking Therapeutic Potential with Antibodies
ProSci brand brings more than 25 years of antibody expertise in developing high quality single domain Variable Heavy domain of Heavy chain (VHH), recombinant, monoclonal, and polyclonal antibodies. In addition, our custom antibody services provide the flexibility to tailor antibody development and production for almost any need. This white paper provides examples of the use of ProSci antibodies for the study of programmed cell death.
Many life forms rely on a process known as programmed cell death. Programmed cell death is essential for the proper functioning of our bodies, and research on this topic has shown us how to better understand and control it.
Many studies that utilized ProSci antibodies have been published in high-impact journals. In the paper published in the journal Immunity (2013) titled “The TACI receptor regulates T-cell-independent marginal zone B cell responses through innate activation-induced cell death,” Figgett et al. reported their findings that TLR4 activation by LPS induces MZ B cell death via TACI-dependent upregulation of Fas and FasL expression, which is impaired in Tnfrsf13b mice. The ProSci Bcl-s antibody was used in this study for western blot analysis (1). Mohmood et al. published a paper in the journal Nature Communication in 2021 titled “β-actin dependent chromatin remodeling mediates compartment level changes in 3D genome architecture.” In this paper, the researchers studied the role of ß-actin in chromatin accessibility and gene expression. This study used the ProSci GAPDH antibody as a control for western blot (2). In the paper published in the journal EMBO (2004) by Gottfried et al. titled “The mitochondrial ARTS protein promotes apoptosis through targeting XIAP,” the authors investigated the role of ARTS in promoting apoptosis through its targeting of XIAP and finds that ARTS can bind directly to XIAP. The ProSci ARTS/ARTSmGTP/H5 were used for this study for western blot analysis (3).
Overall, these publications have had a major impact on the broader understanding of programmed cell death and how it can be utilized for medical purposes. This knowledge has been used to develop new treatments for cancer and other diseases as well as to better understand how our bodies work and how diseases can be prevented or treated more effectively.
Overview
The purpose of studying apoptosis, a specific type of programmed cell death, is to enhance our understanding of its potential uses in treating diseases like cancer. Given its substantial role in the pathogenesis of various diseases, grasping the core mechanisms of apoptosis is critically important. For instance, in cancer scenarios, the balance between cell division and cell death becomes disrupted, leading to cells no longer receiving signals to initiate apoptosis (4). Chemotherapy drugs and radiation are commonly used treatments that induce programmed cell death or apoptosis in cancer cells.
However, a problem with these treatments is their inability to differentiate between healthy and damaged cells, resulting in healthy cells also being killed during treatment (5, 6). To address this issue, scientists are actively working on developing new medications that can reactivate cellular sensors and initiate the process of apoptosis. The goal is to develop more efficient treatments that can minimize collateral damage to healthy cells.
Research is also underway to deepen our comprehension of necroptosis, an alternative form of programmed cell death, and its effects on the body’s reactions to damage and infection (7). Another fascinating avenue of inquiry revolves around autophagy, yet another variant of programmed cell death. Its possible contributions towards the creation of new treatments for age-associated disorders like Alzheimer’s and Parkinson’s disease are currently under scrutiny. Intriguingly, it’s been observed that faulty autophagy mechanisms are a recurring theme among neurodegenerative diseases, a group that also encompasses Alzheimer’s and Parkinson’s disease (8, 9).
In the field of programmed cell death research, antibodies are recognized as indispensable tools. These tools allow scientists to identify and quantify proteins implicated in programmed cell death, along with exploring the related routes and mechanisms.
Furthermore, their role is of utmost importance in determining the impact of various drugs or therapies on programmed cell death. Through the utilization of antibodies, scientists can acquire valuable knowledge regarding the mechanisms underlying programmed cell death, as well as investigate potential approaches to modulate or suppress these mechanisms. Consequently, this offers potential avenues for the management of diseases.
Through a multitude of investigations, researchers are gaining a better understanding of the complex function of programmed cell death that maintains healthy tissue and facilitates favorable survival outcomes.
In this white paper, we will take a closer look at the various types of programmed cell death and how each can be applied to the advancement of medical and scientific research as well as innovative patient therapies.
Exploring Programmed Cell Death: A New Frontier in Disease Treatment
Cell death is a critical biological process that results from activating an intracellular program brought on by various stimuli, such as cellular stress or DNA damage. It typically manifests via two primary mechanisms: apoptosis, initiated by internal signals, and necrosis, which can be triggered by excessive external stimuli.
Apoptosis, often called “programmed cell death,” is an innate response of a cell to certain stimuli, predominantly damage or disease. This is a tightly controlled process characterized by morphological transformations and enzymatic biochemical reactions that remove the cell from the body with minimal disturbance to the surrounding tissue (10, 11). This process is vital in maintaining proper tissue homeostasis. Disruptions in the apoptotic process can lead to various human ailments, such as autoimmune diseases, neurodegenerative disorders, ischemic damage, and cancer (12).
Conversely, necrosis is commonly characterized as a form of cell death that is not under precise biological regulation. This process typically arises from severe damage to the cellular membrane, leading to the leakage of cellular contents. This leakage often initiates an inflammatory reaction that causes damage to surrounding tissues (10, 11). Necrosis can coincide with apoptosis, depending on various factors, such as the intensity and duration of the triggering stimulus and the availability of specific enzymes called caspases (12).
Programmed cell death exists in various distinctive forms, including autophagy, necroptosis, ferroptosis, pyroptosis, parthanatos, and erebosis, a recently uncovered mechanism specific to Drosophila. Unique characteristics and biological functions differentiate the various modes of cell death. Diverse forms of cell death may either promote cell survival or lead to the death of a cell, depending on the environment and signals that trigger the process. Understanding the diverse mechanisms and interactions for these types of cell death is crucial in creating innovative treatment approaches for cancer and other diseases.
The Intrinsic and Extrinsic Pathways of Apoptosis
Apoptosis can occur via two main pathways: the extrinsic and intrinsic pathways. The extrinsic pathway of apoptosis refers to the process through which cells activate programmed cell death in response to external stimuli, such as signals originating from adjacent cells or the immune system. This pathway’s activation is initiated by binding specific ligands to death receptors located on the cell surface, including but not limited to tumor necrosis factor (TNF) or Fas ligand (13). Based on the triggering stimulus and nature of the components involved, at least two apoptotic pathways can be differentiated: one involving receptor systems and one triggered by cytotoxic stress (14).
The extrinsic apoptosis pathway can be initiated through the death receptors, such as Fas, TNF receptor (TNFR), and TNF-related apoptosis-inducing ligand (TRAIL) receptor. The binding of these receptors to their ligands activates caspases, leading to apoptotic cell death (13, 14). Cytotoxic stress can also induce apoptosis. Chemotherapeutic drugs can induce apoptosis and the upregulation of death ligands or their receptors. Downstream events following cytotoxic stress-induced DNA damage and the signaling pathways that lead to the induction of apoptosis may be either dependent or independent of death receptor signaling (15).
The intrinsic apoptosis pathway, also called the mitochondrial pathway, is triggered by various forms of cellular stress, such as DNA damage, oxidative stress, and growth factor deprivation. The pathway is regulated by the BCL-2 protein family, which includes members that exhibit both pro-apoptotic and anti-apoptotic properties. In response to cellular stress, pro-apoptotic proteins, such as Bax and Bak, are activated and undergo oligomerization. This process leads to the formation of pores in the outer mitochondrial membrane.
This process leads to the release of cytochrome c from the mitochondria and its subsequent translocation to the cytosol. Cytochrome c protein binds with Apaf-1, a cytoplasmic protein that initiates apoptosis. This binding results in the formation of an oligomeric apoptosome, then triggering the activation of caspases, which are a family of protease enzymes that play a central role in regulating cell death and inflammation. Apoptosome specifically activates caspase-9, and that, in turn, initiates the activation of downstream effector caspases, such as caspase-3, which leads to the execution phase of apoptosis (14,15).
Key Proteins in the Mechanism of Programmed Cell Death
Programmed cell death involves several key proteins in its regulatory pathways. These proteins play a pivotal role in the initiation, execution, and regulation of apoptosis, coordinating cellular events leading to controlled cell death. The intricate balance between pro-apoptotic and anti-apoptotic proteins ensures proper cell function and organismal health.
Caspases: Caspases, short for cysteine-aspartic proteases, are a family of proteins that play vital roles in programmed cell death, proliferation, and inflammation (16). These proteins are synthesized as inactive precursors and are activated by specific stimuli or signals that initiate a cascade of events and lead to their activation (17). Once activated, they can inactivate or activate substrates, triggering a cascade of signaling events that can lead to an imbalance between cell survival and cell death (18, 19, 20).
Caspase activation and function is a complex process regulated by numerous signals and pathways within cells ranging from interactions with death receptors on the cell surface to internal cellular stress signals and immune responses to interventions from therapeutic treatments (21).
In mammals, three main routes activate the caspases leading to apoptosis:
- Extrinsic or Death Receptor route: Death ligands (including FasL, TNF-alpha, or TRAIL) bind to specific death receptors on the cell surface to initiate this route. Initiator caspases, including caspase-2, caspase-8, caspase-9, and caspase-10, are recruited and activated through a series of intracellular events. These events lead to apoptosis by triggering the activation of executioner caspases, namely caspase-3, caspase-6, and caspase-7 (21).
- Intrinsic or Mitochondrial route: Intracellular stress signals, including DNA damage, oxidative stress, or growth factor deprivation, initiate this route. These signals alter the mitochondrial membrane potential, leading to the release of cytochrome c into the cytoplasm. Released cytochrome c associates with Apoptotic protease activating factor 1 (Apaf-1) and pro-caspase-9 to form the apoptosome, activating caspase-9. Caspase-9 activation triggers the executioner caspases, which in turn triggers apoptosis (21, 22).
- The cytotoxic lymphocyte initiated Granzyme B route: Immune responses cause the activation of this pathway. Perforin and granzymes are released into target cells by cytotoxic T cells and natural killer (NK) cells. Caspases can be directly activated by Granzyme B, resulting in apoptosis (21).
Granzyme B (GrB): A serine protease found in natural killer cells and cytotoxic T cells activates caspases to induce apoptosis in target cells. GrB can directly cleave and activate caspase-3 or other effector caspases. It can also indirectly activate caspase cascade through Bid, a pro-apoptotic member of the Bcl-2 family. This leads to mitochondrial damage and activation of caspase-9, activating effector caspases like caspase-3 (23).
Granzyme A (GzmA): Granzyme A is a structurally related serine protease essential for natural killer cell-mediated and perf-facilitated tumor control. It induces caspase-independent mitochondrial damage, a required first step for apoptosis, and is also responsible for nuclear lamins and histone H1 cleavage.
Granzyme A can target extracellular and intracellular proteins, such as cytokines, matrix proteins, thrombin receptors, and mitochondrial inner membrane potential. Additionally, it has been reported to generate single-stranded DNA nicks rather than the oligonucleosomal DNA fragments typical of granzyme B-induced apoptosis. The role of granzyme A in tumor clearance is controversial, but it is a critical player in cytotoxicity and cell death (16).
Perforin: Perforin is a protein that plays a role in inducing apoptosis in target cells. It is released by cytotoxic T lymphocytes and natural killer cells and forms pores in the target cell membrane, allowing granzymes to enter the cell and initiate apoptosis. Perforin is vital for cytotoxic effector function and is indispensable in granzyme-mediated apoptosis (24).
Bcl-2 family: The Bcl-2 family is a protein group that controls cell death. The presence of conserved sequence motifs characterizes these proteins called Bcl-2 homology motifs and a transmembrane region that forms the interaction sites and determines the intracellular location.
The Bcl-2 family comprises both pro-survival and pro-apoptotic members. Pro-survival members of the family, such as Bcl-2 and Bcl-xL, interact with pro-apoptotic members, such as Bax and Bak, to prevent the release of cytochrome c from the mitochondria. This inhibition of cytochrome c release inhibits apoptosome assembly and the subsequent activation of caspases, thus preventing apoptosis (25, 26).
The pro-apoptotic member of proteins contains a BH3 motif, which binds to pro-survival Bcl-2 proteins and prevents them from inhibiting apoptosis. This binding is necessary for the activation of the caspase. In addition, pro-apoptotic Bcl-2 proteins can also bind to autophagy regulators, such as Beclin-1, to induce autophagy, another type of programmed cell death. Viruses have also taken advantage of the Bcl-2 family by encoding their own Bcl-2 proteins, some of which act as apoptosis inhibitors (26).
Fas/FasL: Fas and Fas ligands are crucial in regulating apoptosis. The Fas receptor, also known as CD95 and tumor necrosis factor receptor superfamily member, is located on the surface of various types of cells. It interacts with its corresponding ligand, FasL (also called FasL/CD95L), to initiate a signal transduction pathway that leads to apoptosis. The interaction between FasL and the Fas receptor initiates a series of events that lead to the recruitment and activation of apoptosis-initiating proteases, such as caspase-8 and caspase-10. These proteases subsequently induce apoptosis through various molecules (27).
Inhibitors of Apoptosis Proteins (IAPs): IAPs act as modulators of apoptosis by inhibiting the activity of caspases. This makes them essential in determining cell survival and death, therefore suggesting potential for therapeutic manipulation (25). Overexpression of IAP proteins is often seen in cancers, making them potential targets for therapeutic intervention (28).
Various Types of Programmed Cell Death
Over the last few decades, various regulated cell death pathways have been identified, each relying on a different subset of proteins for their activation and execution. Apoptosis, necroptosis, and pyroptosis are the most-understood forms of cell death. Still, other pathways are being discovered: autophagy-dependent cell death (ADCD), ferroptosis, parthanatos, and, more recently, a Drosophila-specific cell death mechanism called erebosis.
These different forms of cell death are interconnected and share components with other pathways. Studying cell death pathways and their interconnectivity is important for understanding the physiological and mechanistic aspects of these signaling pathways, leading to better disease treatments caused by deregulated or dysfunctional cell death signaling.
Necroptosis: Necroptosis is a type of controlled cell death resembling necrosis, which causes cells to expand and discharge cytoplasmic material (7). The receptor-interacting protein kinases (RIPKs) RIPK1, RIPK3, and Mixed Lineage Kinase Domain-Like Protein (MLKL) are all activated during necroptosis, which is caspase-independent (29).
Various factors, including cancer, viral infections, and tissue damage, can bring it on. It is characterized by cell swelling, membrane rupture, and the release of intracellular contents, including damage-associated molecular patterns (DAMPs) that can draw immune cells.
Necroptosis has been demonstrated to affect cancer prognosis; some researchers suggest that it suppresses tumor growth, while others claim it encourages tumor growth. Additionally, the production of cytokines during necroptosis might cause inflammation and further encourage tumor growth (29).
Pyroptosis: Pyroptosis differs from other cell death types. It is characterized by cell swelling, pore formation on cell membranes, rupture and bubbling of plasma membranes, moderate chromatin condensation, gasdermin (GSDM) cleavage, formation of inflammasomes, and IL-18 and IL-1β release (30).
Intracellular pathogens primarily trigger pyroptosis and facilitate rapid clearance of various infections by removing intracellular replication niches and boosting the host’s defense response (31). It is regulated by inflammasome-associated caspases, such as CASP1, CASP4, CASP5, and CASP11 (mouse), while some caspases associated with apoptosis, such as CASP3 and CASP8 also play a role in pyroptosis (27). It is a crucial mechanism in protecting the host against intracellular pathogens and contributes to antitumor immunity. The induction of pyroptosis can induce potent antitumor activity (29).
Autophagy-dependent cell death (ADCD): Autophagy-dependent cell death (ADCD) is a form of caspase-independent programmed cell death. It’s characterized by an over-activation of autophagy, a cellular degradation process used by eukaryotic cells for maintaining homeostasis and managing lipid metabolism (29).
This over-activation leads to a high degree of self-digestion of cellular material in autolysosomes, moving past the threshold of cell survival24. Autophagy substrates (DAMPs) and pathogen-associated molecular pattern (PAMPs) molecules trigger innate immunity whose elimination through autophagy is necessary for immune homeostasis to protect the cells from exposed membranes and other organelles (29). ADCD occurs during the response of certain cancer cells to specific drugs or natural compounds, showing its relevance in biological and pathological contexts (29, 32).
Ferroptosis: Ferroptosis is regulated cell death dependent on iron and excessive lipid peroxidation, leading to oxidative membrane damage and ruptured plasma membranes. It is distinct from other forms of cell death, regulated by multiple metabolic events, including redox balance, iron handling, and mitochondrial activity (33).
Iron accumulation and lipid peroxidation are the two fundamental processes that trigger ferroptosis (27). In combination with immunotherapy, targeted therapies (inducers or inhibitors) against ferroptosis may exert potent antitumor activity, even in tumors resistant to immune checkpoint inhibitors (29).
Parthanatos: Parthanatos is a form of programmed cell death initiated by PAR polymer, a component of Poly-ADP-ribose polymerase-1 (PARP-1) and is distinct from other cell death processes such as necrosis and apoptosis. It is caused by the accumulation of Poly (ADP ribose) (PAR) and the nuclear translocation of apoptosis-inducing factor (AIF) from mitochondria. Parthanatos’ main mechanism of action is the production of PAR polymer, which leads to NAD+ depletion and the release of apoptosis-inducing factor (AIF), ultimately resulting in cell death.
Other proposed mechanisms of action of parthanatos include mitochondrial permeability transition (PT) and the loss of mitochondrial membrane potential. Understanding parthanatos will help in expanding therapeutic options for PARP-1-related diseases (34).
Erebosis: Erebosis is a newly identified type of cell death discovered in the gut enterocytes of adult Drosophila, or fruit flies. This process differs from known cell death pathways, such as apoptosis, necrosis, or autophagy. It is characterized by the loss of the cytoskeleton, cell adhesion, and organelles. Enterocytes surrounding the erebotic cells often protrude towards them, and old differentiated cells are believed to typically die by apoptosis. Further investigation will be conducted to determine the molecular mechanisms by which erebosis occurs and its role in gut tissue homeostasis (35).
Summary: Exploring the Research on Programmed Cell Death
The exploration of programmed cell death holds profound potential for advancing science and medicine. This fundamental biological event can shed light on diverse domains, including cancer therapy, stem cell research, regenerative medicine, understanding disease mechanisms, drug development, and fundamental biological understanding. Broadly, the course of programmed cell death research promises to enrich our knowledge about the intricacies of cellular life and death, thereby creating opportunities for innovative disease therapies.
Below is a sample of ProSci antibodies related to Apoptosis research. Visit Prosci website to find the right antibodies to help with your research: https://www.prosci-inc.com
| Capases | |
| Cat # | Product |
| 1128 | Caspase-10 Antibody |
| 2071 | Caspase-9 Antibody |
| PM-2015 | Apaf-1 Antibody [2E10] |
| Necroptosis | |
| Cat # | Product |
| 2283 | RIP3 Antibody |
| 3661 | TRAF2 Antibody |
| 3291 | RIPK1 Antibody |
| Ferroptosis | |
| Cat # | Product |
| 14-907 | GPX4 Antibody |
| 13-240 | NRF2 Antibody |
| 16-557 | HSPB1 Antibody, KO Validated |
| Serine Proteases | |
| Cat # | Product |
| 18-708 | GZMB Antibody |
| 22-069 | GZMA Antibody |
| 13-029 | Perforin Antibody |
| Pyroptosis | |
| Cat # | Product |
| 5447 | NALP3 Antibody |
| 16-910 | GSDMD Antibody |
| 3463 | Caspase-1 Antibody |
| Parthantos | |
| Cat # | Product |
| 13-327 | PARP1 Antibody, KO Validated |
| 2267 | AIF Antibody |
| 15-262 | MIF Antibody |
| BCL-2 | |
| Cat # | Product |
| 3343 | BAD Antibody |
| 3335 | Bcl-2 Antibody |
| 3351 | Bax Antibody |
References:
- Figgett WA, Fairfax K, Vincent FB, Le Page MA, Katik I, Deliyanti D, Quah PS, Verma P, Grumont R, Gerondakis S, Hertzog P, O’Reilly LA, Strasser A, Mackay F. The TACI receptor regulates T-cell-independent marginal zone B cell responses through innate activation-induced cell death. Immunity. 2013 Sep 19;39(3):573-83. doi: 10.1016/j.immuni.2013.05.019. Epub 2013 Sep 5. PMID: 24012421.
- Mahmood SR, Xie X, Hosny El Said N, Venit T, Gunsalus KC, Percipalle P. β-actin dependent chromatin remodeling mediates compartment level changes in 3D genome architecture. Nat Commun. 2021 Sep 2;12(1):5240. doi: 10.1038/s41467-021-25596-2. PMID: 34475390; PMCID: PMC8413440.
- Gottfried Y, Rotem A, Lotan R, Steller H, Larisch S. The mitochondrial ARTS protein promotes apoptosis through targeting XIAP. EMBO J. 2004 Apr 7;23(7):1627-35. doi: 10.1038/sj.emboj.7600155. Epub 2004 Mar 18. PMID: 15029247; PMCID: PMC391065
- Wong RS. Apoptosis in cancer: from pathogenesis to treatment. J Exp Clin Cancer Res. 2011 Sep 26;30(1):87. doi: 10.1186/1756-9966-30-87. PMID: 21943236; PMCID: PMC3197541
- Zhao T, He Q, Xie S, Zhan H, Jiang C, Lin S, Liu F, Wang C, Chen G, Zeng H. A novel Mcl-1 inhibitor synergizes with venetoclax to induce apoptosis in cancer cells. Mol Med. 2023 Jan 19;29(1):10. doi: 10.1186/s10020-022-00565-7. PMID: 36658493; PMCID: PMC9854187.
- Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020 Jul;17(7):395-417. doi: 10.1038/s41571-020-0341-y. Epub 2020 Mar 23. PMID: 32203277; PMCID: PMC8211386.
- Initiation and execution mechanisms of necroptosis: an overview. Cell Death Differ. 2017 Jul;24(7):1184-1195. doi: 10.1038/cdd.2017.65.
- Liu J, Li L. Targeting Autophagy for the Treatment of Alzheimer’s Disease: Challenges and Opportunities. Front Mol Neurosci. 2019 Aug 22;12:203. doi: 10.3389/fnmol.2019.00203. PMID: 31507373; PMCID: PMC6713911.
- Rai SN, Tiwari N, Singh P, Mishra D, Singh AK, Hooshmandi E, Vamanu E, Singh MP. Therapeutic Potential of Vital Transcription Factors in Alzheimer’s and Parkinson’s Disease With Particular Emphasis on Transcription Factor EB Mediated Autophagy. Front Neurosci. 2021 Dec 14;15:777347. doi: 10.3389/fnins.2021.777347. PMID: 34970114; PMCID: PMC8712758.
- Erebosis is a new type of cell death for tissue homeostasis in the Drosophila intestine. PLoS Biol 20(4): e3001614. doi.org/10.1371/journal.pbio.3001614
- The concept of intrinsic versus extrinsic apoptosis. Biochem J. 2022 Feb 11;479(3):357-384. doi: 10.1042/BCJ20210854.
- The proteins and the mechanisms of apoptosis: a mini-review of the fundamentals. Hippokratia. 2007 Jul;11(3):108-13. PMID: 19582203
- Green R, Llambi F. Cell Death Signaling. Cold Spring Harb Perspect Biol. 2015 Dec 1;7(12):a006080. doi: 10.1101/cshperspect.a006080. PMID: 26626938; PMCID: PMC4665079.
- Bots M, Medema JP. Granzymes at a glance. J Cell Sci. 2006 Dec 15;119(Pt 24):5011-4. doi: 10.1242/jcs.03239. PMID: 17158907.
- Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol. 2002 Oct;2(10):735-47. doi: 10.1038/nri911. PMID: 12360212.
- Bots M, Medema JP. Granzymes at a glance. J Cell Sci. 2006 Dec 15;119(Pt 24):5011-4. doi: 10.1242/jcs.03239. PMID: 17158907
- Autophagy, ferroptosis, pyroptosis, and necroptosis in tumor immunotherapy. Signal Transduct Target Ther. 2022 Jun 20;7(1):196. doi: 10.1038/s41392-022-01046-3
- Targeting IAP (inhibitor of apoptosis) proteins for therapeutic intervention in tumors. Curr Cancer Drug Targets. 2008 Mar;8(2):110-7. doi: 10.2174/156800908783769373.
- Caspase activation cascades in apoptosis. Biochem Soc Trans. 2008 Feb;36(Pt 1):1-9. doi: 10.1042/BST0360001.
- Caspase activation pathways: some recent progress. Cell Death Differ. 2009 Jul;16(7):935-8. doi: 10.1038/cdd.2009.59.
- Human caspases: activation, specificity, and regulation. J Biol Chem. 2009 Aug 14;284(33):21777-21781. doi: 10.1074/jbc.R800084200.
- Cell pyroptosis in health and inflammatory diseases. Cell Death Discov. 2022 Apr 11;8(1):191. doi: 10.1038/s41420-022-00998-3.
- Apoptotic and non-apoptotic roles of caspases in neuronal physiology and pathophysiology. Nat Rev Neurosci. 2012 May 18;13(6):395-406. doi: 10.1038/nrn3228.
- Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol. 2002 Oct;2(10):735-47. doi: 10.1038/nri911. PMID: 12360212
- Banjara S, Suraweera CD, Hinds MG, Kvansakul M. The Bcl-2 Family: Ancient Origins, Conserved Structures, and Divergent Mechanisms. Biomolecules. 2020 Jan 12;10(1):128. doi: 10.3390/biom10010128. PMID: 31940915; PMCID: PMC7022251.
- Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol. 2008 Jan;9(1):47-59. doi: 10.1038/nrm2308. PMID: 18097445
- Yu WR, Fehlings MG. Fas/FasL-mediated apoptosis and inflammation are key features of acute human spinal cord injury: implications for translational, clinical application. Acta
- Targeting IAP (inhibitor of apoptosis) proteins for therapeutic intervention in tumors. Curr Cancer Drug Targets. 2008 Mar;8(2):110-7. doi: 10.2174/156800908783769373.
- Research progresses of molecular mechanism of pyroptosis and its related diseases. Immunobiology. 2020 Mar;225(2):151884. doi: 10.1016/j.imbio.2019.11.019.
- Cell pyroptosis in health and inflammatory diseases. Cell Death Discov. 2022 Apr 11;8(1):191. doi: 10.1038/s41420-022-00998-3.
- Research progresses of molecular mechanism of pyroptosis and its related diseases. Immunobiology. 2020 Mar;225(2):151884. doi: 10.1016/j.imbio.2019.11.019.
- Organelle-specific mechanisms of drug-induced autophagy-dependent cell death. Matrix Biol. 2021 Jun;100-101:54-64. doi: 10.1016/j.matbio.2020.12.003
- Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021 Apr;22(4):266-282. doi: 10.1038/s41580-020-00324-8
- David KK, Andrabi SA, Dawson TM, Dawson VL. Parthanatos, a messenger of death. Front Biosci (Landmark Ed). 2009 Jan 1;14(3):1116-28. doi: 10.2741/3297. PMID: 19273119; PMCID: PMC4450718.
- Erebosis is a new type of cell death for tissue homeostasis in the Drosophila intestine. PLoS Biol 20(4): e3001614. doi.org/10.1371/journal.pbio.3001614
Roles of RIPKs in Immune Responses, Inflammation, and Cell Death
Exploring the Roles of RIPKs in Immune Responses, Inflammation, and Cell Death: Recent Findings
Introduction
The RIPK family comprises seven proteins (RIP1, RIP3, RIP4, RIP5, RIP6, RIP7, and DIK) that are involved in cytokine stimulation, pathogen infection, DNA damage, and inflammation signaling pathways. They help to initiate various responses, from immune cell activation to cell death. They are also essential sensors of intracellular and extracellular stresses, regulating programmed necrosis, apoptosis, and other cell death pathways. RIPKs are critical for proper immune function and modulating the NF-kB pathway and cell death program (1,2,3).
Genesee Scientific’s ProSci portfolio brings more than 25 years of antibody expertise in developing high quality single domain Variable Heavy domain of Heavy chain (VHH), recombinant, monoclonal, and polyclonal antibodies. In addition, Prosci custom antibody services provide the flexibility to tailor antibody development and production for almost any need. This white paper provides examples of the use of ProSci RIPK antibodies and highlights recent findings on the roles of RIPKs in signaling pathways involved in immune responses, inflammation, and cell death.
“RIPK3-ZBP1 Interaction Facilitated by Caspase-6: Essential Role in Inflammasome Activation and Host Defense Against Influenza A Virus”
The paper published in 2020 by Zheng et al. titled “Caspase-6 Is a Key Regulator of Innate Immunity, Inflammasome Activation, and Host Defense” demonstrated that caspase-6 modulates innate immunity, inflammasome activation, and multiple cell death pathways (4).
Caspases are a family of cysteine proteases essential in regulating various biological processes such as host cell death, innate immune responses, and homeostasis (5). Caspases are categorized into inflammatory caspases (caspase-1, 4, 5, 11, and 12) and apoptotic caspases (caspase-2, 3, 6, 7, 8, 9, and 10). The apoptotic caspases can be further categorized into the initiator caspases (caspase-2, 8, 9, and 10) and the effector (or executioner) caspases (caspase-3, 6, and 7) (6). The significance of caspase-6 in innate immunity remains an essential focus of the investigation, as its precise mechanisms in regulating cell death, inflammasome activation, and host defense mechanisms are still not fully understood.
Caspase-6 plays a crucial role in defending the host against Influenza A virus (IAV) infection. To examine the impact of caspase-6 deficiency, both Casp6+/− and Casp6−/− littermate mice were infected with IAV via the intranasal route. The findings revealed that a higher number of Casp6−/− mice succumbed to IAV infection. The authors conducted further investigations to understand the involvement of caspase-6 in cell death pathways induced by IAV, including pyroptosis, apoptosis, and necroptosis. They observed that Casp6−/− mice exhibited decreased levels of IL-1β in bronchoalveolar lavage fluid (BALF) on day seven post-infection and a significant increase in viral titer in their lungs compared to wild-type (WT) mice.
Interestingly, caspase-6 appears to function independently of priming, STAT1 activation, IFN signaling, and ZBP1 expression. Moreover, the results demonstrated that caspase-6 is not essential for canonical NLRP3 inflammasome activation, NLRC4 inflammasome activation, AIM2 inflammasome activation, or PYRIN inflammasome activation in response to pathogen infections. The CRISPR gene editing technique was employed to knockout caspase-6 in mouse embryonic fibroblasts (MEFs) to validate these findings at the cellular level further. The researchers observed reduced cell death following IAV infection compared to WT MEFs.
Furthermore, WT and caspase-6-deficient bone marrow-derived macrophages (BMDMs) were stimulated with ATP or nigericin or infected with various bacteria and viruses. It was found that the reintroduction of caspase-6 into Casp6-/- BMDMs led to increased caspase-1 activation induced by IAV infection both in its WT and catalytically dead forms. This suggests that the caspase activity of caspase-6 is not necessary for inflammasome activation.
The ProSci rabbit anti-RIPK3 antibody (ProSci Cat. #: 2283) was used to immunoprecipitate RIPK3 during IAV infection to determine the role of RIPK3 and caspase-8 in caspase-6 activation during IAV infection. Rabbit anti-RIPK3 antibody was also used in co-immunoprecipitation to investigate the interaction between caspase-6 and RIPK3 and their interaction with ZBP1 and RIPK1.
These findings suggest that caspase-6 plays a vital role in regulating multiple cell death pathways in response to DAMPs or PAMPs and protecting the host against IAV infection.
“RIPK3’s Role in Balancing Necrosomal Degradation and Maturation: A New Mechanism Mediated by Triad3a with Implications for Controlling Inflammation and Necroptosis”
The study published in 2018 by Alturki et al. titled “Triad3a induces the degradation of early necrosome to limit RipK1-dependent cytokine production and necroptosis” investigated the role of E3-ubiquitin ligase Triad3a in regulating cell death during infections. They demonstrated that Triad3a plays a key role in regulating cell death during infections by mediating the degradation of RIPK3 interacting proteins and inducing inflammatory cell death, a new mechanism of shutting off necrosome signaling (8).
TLR signaling induces MyD88-dependent and TRIF-dependent signaling, which leads to the production of cytokines and chemokines and the recruitment of myeloid cells (9,10). Necrosis is a pathway of regulated necrosis, called “necroptosis,” and is induced by TNF-R or IFN-IR engagement. Necrosome signaling involves RIPK1-FADD-caspase-8 interaction and leads to the phosphorylation of RIPK3 (11,12). Macrophage-like cells are present throughout the body and play a significant role in initiating inflammatory responses to control pathogens.
The authors investigated the role of RIPK1 and RIPK3 in necrosome signaling and the impact of deubiquitinating enzyme CLYD and ubiquitin-editing enzyme A20 knockdown on the degradation of RIPK1. It was found that an inhibitor of necroptosis necrostatin 1 (Nec-1) completely nullified the phosphorylation of RIPK3, degradation of RIPK1, and cell death by necroptosis. Loss of RIPK3 or mixed lineage kinase domain-like pseudokinase (MLKL) led to a decrease in RIPK3 phosphorylation and protection against necroptosis. CYLD deficiency had little effect on RIPK1 degradation but led to decreased phosphorylation of RIPK3 and necroptosis. Stimulation of cells with lipopolysaccharide (LPS) and pan-caspase inhibitor zVAD had a marginal impact on cell death at earlier points in time.
Co-immunoprecipitation with an anti-RIPK3 antibody showed that Nec-1 completely abrogated the phosphorylation of RIPK3, degradation of RIPK1, and cell death by necroptosis. The combined stimulation of toll-like receptor 4 (TLR4) by LPS and caspase inhibition by zVAD caused the phosphorylation of RIPK1 and RIPK3. Caspase-8 inhibitor zIETD failed to induce the loss of RIPK1 expression or necroptosis of macrophages. K48-ubiquitination of RIPK1 and caspase-8 following necrosome activation was significantly reduced in RIPK3-deficient macrophages. In macrophages, CYLD and A20 knockdown enhanced the degradation of RIPK1. The results revealed that early degradation of the necrosome was a mechanism of cellular escape from necroptosis and cytokine expression. Knockdown of Triad3a resulted in reduced degradation of RIPK1, FADD, and caspase-8 and enhanced the expression.
In this study, ProSci’s rabbit anti-RIPK3 antibody (ProSci Cat. #: 2283) antibody was used for western blot and immunoprecipitation to investigate its role in the degradation of necrosomal proteins during necroptosis.
This study investigated a new mechanism of self-regulation built into necrosome signaling, which involves the expression and phosphorylation of CYLD. The results show that forming an early necrosome platform triggers a decision between degradation via K48-ubiquitination or cell death via RIPK3 kinase activity and MLKL recruitment. The study also found that the kinase function of RIPK3 is necessary to activate necroptosis, but its interaction with RIPK3 leads to the degradation of RIPK1 and other necrosomal proteins. Finally, applying small molecule inhibitors of RIPK3 can prevent activation of the late necrosome, leading to cells that are resistant to necroptotic challenges. The study reveals that necrosome signaling is regulated by a complex mechanism involving multiple kinases and ubiquitin ligases.
Conclusion
This white paper presents studies that underscore the crucial role played by the RIPK family—specifically RIPK3—in regulating immune responses, inflammation, and cell death processes.
Zheng et al. highlight how caspase-6 has a significant role in activating the NLRP3 inflammasome through ZBP1 during IAV infection. The authors emphasize the importance of caspase-6 activation and how it relies on interactions with RIPK3 and caspase-8. Such findings hold promise for potential therapeutic approaches targeting caspase-6 along with its interplay with RIPK3, ZBP1, and RIPK1 to address IAV infection effectively (4).
In addition, Alturki et al. revealed an innovative mechanism where Triad3a mediates downregulation of the necrosome—a central signaling complex tied to inflammation and necroptosis processes. This process involves degrading key proteins such as RIPK1 and other necrosomal proteins. Balancing the degradation and maturation of the necrosome allows for better control over the extent of necroptotic cell death. Consequently, this Triad3a-dependent mechanism presents novel therapeutic strategies that can be explored to influence inflammatory responses and regulate necroptosis (8).
Overall, these studies contribute to a more comprehensive understanding of the complex role played by the RIPK family in modulating immune responses, inflammation, and cell death processes. The knowledge gained from these studies offers the potential for developing innovative therapeutic interventions targeting various diseases, including those related to viral infections and inflammatory conditions.
Visit Prosci website to learn more about anti-RIPK3 and related antibodies needed to help with your research: https://www.prosci-inc.com
References:
- Zhang D, Lin J, Han J. Receptor-interacting protein (RIP) kinase family. Cell Mol Immunol. 2010 Jul;7(4):243-9. doi: 10.1038/cmi.2010.10. Epub 2010 Apr 12. PMID: 20383176; PMCID: PMC4003224.
- Declercq W, Vanden Berghe T, Vandenabeele P. RIP kinases at the crossroads of cell death and survival. Cell. 2009 Jul 23;138(2):229-32. doi: 10.1016/j.cell.2009.07.006. PMID: 19632174.
- Dondelinger Y, Jouan-Lanhouet S, Divert T, Theatre E, Bertin J, Gough PJ, Giansanti P, Heck AJ, Dejardin E, Vandenabeele P, Bertrand MJ. NF-κB-Independent Role of IKKα/IKKβ in Preventing RIPK1 Kinase-Dependent Apoptotic and Necroptotic Cell Death during TNF Signaling. Mol Cell. 2015 Oct 1;60(1):63-76. doi: 10.1016/j.molcel.2015.07.032. Epub 2015 Sep 3. PMID: 26344099.
- Zheng M, Karki R, Vogel P, Kanneganti TD. Caspase-6 Is a Key Regulator of Innate Immunity, Inflammasome Activation, and Host Defense. Cell. 2020 Apr 30;181(3):674-687.e13. doi: 10.1016/j.cell.2020.03.040. Epub 2020 Apr 15. PMID: 32298652; PMCID: PMC7425208.
- Van Opdenbosch N, Lamkanfi M. Caspases in Cell Death, Inflammation, and Disease. Immunity. 2019 Jun 18;50(6):1352-1364. doi: 10.1016/j.immuni.2019.05.020. PMID: 31216460; PMCID: PMC6611727.
- Man SM, Kanneganti TD. Converging roles of caspases in inflammasome activation, cell death and innate immunity. Nat Rev Immunol. 2016 Jan;16(1):7-21. doi: 10.1038/nri.2015.7. Epub 2015 Dec 14. PMID: 26655628; PMCID: PMC4915362.
- Kuriakose T, Man SM, Malireddi RK, Karki R, Kesavardhana S, Place DE, Neale G, Vogel P, Kanneganti TD. ZBP1/DAI is an innate sensor of influenza virus triggering the NLRP3 inflammasome and programmed cell death pathways. Sci Immunol. 2016 Aug 5;1(2):aag2045. doi: 10.1126/sciimmunol.aag2045. Epub 2016 Aug 12. PMID: 27917412; PMCID: PMC5131924.
- Alturki NA, McComb S, Ariana A, Rijal D, Korneluk RG, Sun SC, Alnemri E, Sad S. Triad3a induces the degradation of early necrosome to limit RipK1-dependent cytokine production and necroptosis. Cell Death Dis. 2018 May 22;9(6):592. doi: 10.1038/s41419-018-0672-0. PMID: 29789521; PMCID: PMC5964080.
- Akira S. TLR signaling. Curr Top Microbiol Immunol. 2006;311:1-16. doi: 10.1007/3-540-32636-7_1. PMID: 17048703.
- Eckmann L, Kagnoff MF. Cytokines in host defense against Salmonella. Microbes Infect. 2001 Nov-Dec;3(14-15):1191-200. doi: 10.1016/s1286-4579(01)01479-4. PMID: 11755407.
- He S, Wang L, Miao L, Wang T, Du F, Zhao L, Wang X. Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-alpha. Cell. 2009 Jun 12;137(6):1100-11. doi: 10.1016/j.cell.2009.05.021. PMID: 19524512.
- Christofferson DE, Yuan J. Necroptosis as an alternative form of programmed cell death. Curr Opin Cell Biol. 2010 Apr;22(2):263-8. doi: 10.1016/j.ceb.2009.12.003. Epub 2010 Jan 4. PMID: 20045303; PMCID: PMC2854308.
Hidden Power of Programmed Cell Death
A Deep Dive into the Hidden Power of Programmed Cell Death: Unlocking Therapeutic Potential with Antibodies
Genesee Scientific’s ProSci brand brings more than 25 years of antibody expertise in developing high quality single domain Variable Heavy domain of Heavy chain (VHH), recombinant, monoclonal, and polyclonal antibodies. In addition, Proci custom antibody services provide the flexibility to tailor antibody development and production for almost any need. This white paper provides examples of the use of ProSci antibodies for the study of programmed cell death.
Many life forms rely on a process known as programmed cell death. Programmed cell death is essential for the proper functioning of our bodies, and research on this topic has shown us how to better understand and control it.
Many studies that utilized ProSci antibodies have been published in high-impact journals. In the paper published in the journal Immunity (2013) titled “The TACI receptor regulates T-cell-independent marginal zone B cell responses through innate activation-induced cell death,” Figgett et al. reported their findings that TLR4 activation by LPS induces MZ B cell death via TACI-dependent upregulation of Fas and FasL expression, which is impaired in Tnfrsf13b mice. The ProSci Bcl-s antibody was used in this study for western blot analysis (1). Mohmood et al. published a paper in the journal Nature Communication in 2021 titled “β-actin dependent chromatin remodeling mediates compartment level changes in 3D genome architecture.” In this paper, the researchers studied the role of ß-actin in chromatin accessibility and gene expression. This study used the ProSci GAPDH antibody as a control for western blot (2). In the paper published in the journal EMBO (2004) by Gottfried et al. titled “The mitochondrial ARTS protein promotes apoptosis through targeting XIAP,” the authors investigated the role of ARTS in promoting apoptosis through its targeting of XIAP and finds that ARTS can bind directly to XIAP. The ProSci ARTS/ARTSmGTP/H5 were used for this study for western blot analysis (3).
Overall, these publications have had a major impact on the broader understanding of programmed cell death and how it can be utilized for medical purposes. This knowledge has been used to develop new treatments for cancer and other diseases as well as to better understand how our bodies work and how diseases can be prevented or treated more effectively.
Overview
The purpose of studying apoptosis, a specific type of programmed cell death, is to enhance our understanding of its potential uses in treating diseases like cancer. Given its substantial role in the pathogenesis of various diseases, grasping the core mechanisms of apoptosis is critically important. For instance, in cancer scenarios, the balance between cell division and cell death becomes disrupted, leading to cells no longer receiving signals to initiate apoptosis (4). Chemotherapy drugs and radiation are commonly used treatments that induce programmed cell death or apoptosis in cancer cells.
However, a problem with these treatments is their inability to differentiate between healthy and damaged cells, resulting in healthy cells also being killed during treatment (5, 6). To address this issue, scientists are actively working on developing new medications that can reactivate cellular sensors and initiate the process of apoptosis. The goal is to develop more efficient treatments that can minimize collateral damage to healthy cells.
Research is also underway to deepen Prosci comprehension of necroptosis, an alternative form of programmed cell death, and its effects on the body’s reactions to damage and infection (7). Another fascinating avenue of inquiry revolves around autophagy, yet another variant of programmed cell death. Its possible contributions towards the creation of new treatments for age-associated disorders like Alzheimer’s and Parkinson’s disease are currently under scrutiny. Intriguingly, it’s been observed that faulty autophagy mechanisms are a recurring theme among neurodegenerative diseases, a group that also encompasses Alzheimer’s and Parkinson’s disease (8, 9).
In the field of programmed cell death research, antibodies are recognized as indispensable tools. These tools allow scientists to identify and quantify proteins implicated in programmed cell death, along with exploring the related routes and mechanisms.
Furthermore, their role is of utmost importance in determining the impact of various drugs or therapies on programmed cell death. Through the utilization of antibodies, scientists can acquire valuable knowledge regarding the mechanisms underlying programmed cell death, as well as investigate potential approaches to modulate or suppress these mechanisms. Consequently, this offers potential avenues for the management of diseases.
Through a multitude of investigations, researchers are gaining a better understanding of the complex function of programmed cell death that maintains healthy tissue and facilitates favorable survival outcomes.
In this white paper, we will take a closer look at the various types of programmed cell death and how each can be applied to the advancement of medical and scientific research as well as innovative patient therapies. i
Exploring Programmed Cell Death: A New Frontier in Disease Treatment
Cell death is a critical biological process that results from activating an intracellular program brought on by various stimuli, such as cellular stress or DNA damage. It typically manifests via two primary mechanisms: apoptosis, initiated by internal signals, and necrosis, which can be triggered by excessive external stimuli.
Apoptosis, often called “programmed cell death,” is an innate response of a cell to certain stimuli, predominantly damage or disease. This is a tightly controlled process characterized by morphological transformations and enzymatic biochemical reactions that remove the cell from the body with minimal disturbance to the surrounding tissue (10, 11). This process is vital in maintaining proper tissue homeostasis. Disruptions in the apoptotic process can lead to various human ailments, such as autoimmune diseases, neurodegenerative disorders, ischemic damage, and cancer (12).
Conversely, necrosis is commonly characterized as a form of cell death that is not under precise biological regulation. This process typically arises from severe damage to the cellular membrane, leading to the leakage of cellular contents. This leakage often initiates an inflammatory reaction that causes damage to surrounding tissues (10, 11). Necrosis can coincide with apoptosis, depending on various factors, such as the intensity and duration of the triggering stimulus and the availability of specific enzymes called caspases (12).
Programmed cell death exists in various distinctive forms, including autophagy, necroptosis, ferroptosis, pyroptosis, parthanatos, and erebosis, a recently uncovered mechanism specific to Drosophila. Unique characteristics and biological functions differentiate the various modes of cell death. Diverse forms of cell death may either promote cell survival or lead to the death of a cell, depending on the environment and signals that trigger the process. Understanding the diverse mechanisms and interactions for these types of cell death is crucial in creating innovative treatment approaches for cancer and other diseases.
The Intrinsic and Extrinsic Pathways of Apoptosis
Apoptosis can occur via two main pathways: the extrinsic and intrinsic pathways. The extrinsic pathway of apoptosis refers to the process through which cells activate programmed cell death in response to external stimuli, such as signals originating from adjacent cells or the immune system. This pathway’s activation is initiated by binding specific ligands to death receptors located on the cell surface, including but not limited to tumor necrosis factor (TNF) or Fas ligand (13). Based on the triggering stimulus and nature of the components involved, at least two apoptotic pathways can be differentiated: one involving receptor systems and one triggered by cytotoxic stress (14).
The extrinsic apoptosis pathway can be initiated through the death receptors, such as Fas, TNF receptor (TNFR), and TNF-related apoptosis-inducing ligand (TRAIL) receptor. The binding of these receptors to their ligands activates caspases, leading to apoptotic cell death (13, 14). Cytotoxic stress can also induce apoptosis. Chemotherapeutic drugs can induce apoptosis and the upregulation of death ligands or their receptors. Downstream events following cytotoxic stress-induced DNA damage and the signaling pathways that lead to the induction of apoptosis may be either dependent or independent of death receptor signaling (15).
The intrinsic apoptosis pathway, also called the mitochondrial pathway, is triggered by various forms of cellular stress, such as DNA damage, oxidative stress, and growth factor deprivation. The pathway is regulated by the BCL-2 protein family, which includes members that exhibit both pro-apoptotic and anti-apoptotic properties. In response to cellular stress, pro-apoptotic proteins, such as Bax and Bak, are activated and undergo oligomerization. This process leads to the formation of pores in the outer mitochondrial membrane.
This process leads to the release of cytochrome c from the mitochondria and its subsequent translocation to the cytosol. Cytochrome c protein binds with Apaf-1, a cytoplasmic protein that initiates apoptosis. This binding results in the formation of an oligomeric apoptosome, then triggering the activation of caspases, which are a family of protease enzymes that play a central role in regulating cell death and inflammation. Apoptosome specifically activates caspase-9, and that, in turn, initiates the activation of downstream effector caspases, such as caspase-3, which leads to the execution phase of apoptosis (14,15).
Key Proteins in the Mechanism of Programmed Cell Death
Programmed cell death involves several key proteins in its regulatory pathways. These proteins play a pivotal role in the initiation, execution, and regulation of apoptosis, coordinating cellular events leading to controlled cell death. The intricate balance between pro-apoptotic and anti-apoptotic proteins ensures proper cell function and organismal health.
Caspases: Caspases, short for cysteine-aspartic proteases, are a family of proteins that play vital roles in programmed cell death, proliferation, and inflammation (16). These proteins are synthesized as inactive precursors and are activated by specific stimuli or signals that initiate a cascade of events and lead to their activation (17). Once activated, they can inactivate or activate substrates, triggering a cascade of signaling events that can lead to an imbalance between cell survival and cell death (18, 19, 20).
Caspase activation and function is a complex process regulated by numerous signals and pathways within cells ranging from interactions with death receptors on the cell surface to internal cellular stress signals and immune responses to interventions from therapeutic treatments (21).
In mammals, three main routes activate the caspases leading to apoptosis:
- Extrinsic or Death Receptor route: Death ligands (including FasL, TNF-alpha, or TRAIL) bind to specific death receptors on the cell surface to initiate this route. Initiator caspases, including caspase-2, caspase-8, caspase-9, and caspase-10, are recruited and activated through a series of intracellular events. These events lead to apoptosis by triggering the activation of executioner caspases, namely caspase-3, caspase-6, and caspase-7 (21).
- Intrinsic or Mitochondrial route: Intracellular stress signals, including DNA damage, oxidative stress, or growth factor deprivation, initiate this route. These signals alter the mitochondrial membrane potential, leading to the release of cytochrome c into the cytoplasm. Released cytochrome c associates with Apoptotic protease activating factor 1 (Apaf-1) and pro-caspase-9 to form the apoptosome, activating caspase-9. Caspase-9 activation triggers the executioner caspases, which in turn triggers apoptosis (21, 22).
- The cytotoxic lymphocyte initiated Granzyme B route: Immune responses cause the activation of this pathway. Perforin and granzymes are released into target cells by cytotoxic T cells and natural killer (NK) cells. Caspases can be directly activated by Granzyme B, resulting in apoptosis (21).
Granzyme B (GrB): A serine protease found in natural killer cells and cytotoxic T cells activates caspases to induce apoptosis in target cells. GrB can directly cleave and activate caspase-3 or other effector caspases. It can also indirectly activate caspase cascade through Bid, a pro-apoptotic member of the Bcl-2 family. This leads to mitochondrial damage and activation of caspase-9, activating effector caspases like caspase-3 (23).
Granzyme A (GzmA): Granzyme A is a structurally related serine protease essential for natural killer cell-mediated and perf-facilitated tumor control. It induces caspase-independent mitochondrial damage, a required first step for apoptosis, and is also responsible for nuclear lamins and histone H1 cleavage.
Granzyme A can target extracellular and intracellular proteins, such as cytokines, matrix proteins, thrombin receptors, and mitochondrial inner membrane potential. Additionally, it has been reported to generate single-stranded DNA nicks rather than the oligonucleosomal DNA fragments typical of granzyme B-induced apoptosis. The role of granzyme A in tumor clearance is controversial, but it is a critical player in cytotoxicity and cell death (16).
Perforin: Perforin is a protein that plays a role in inducing apoptosis in target cells. It is released by cytotoxic T lymphocytes and natural killer cells and forms pores in the target cell membrane, allowing granzymes to enter the cell and initiate apoptosis. Perforin is vital for cytotoxic effector function and is indispensable in granzyme-mediated apoptosis (24).
Bcl-2 family: The Bcl-2 family is a protein group that controls cell death. The presence of conserved sequence motifs characterizes these proteins called Bcl-2 homology motifs and a transmembrane region that forms the interaction sites and determines the intracellular location.
The Bcl-2 family comprises both pro-survival and pro-apoptotic members. Pro-survival members of the family, such as Bcl-2 and Bcl-xL, interact with pro-apoptotic members, such as Bax and Bak, to prevent the release of cytochrome c from the mitochondria. This inhibition of cytochrome c release inhibits apoptosome assembly and the subsequent activation of caspases, thus preventing apoptosis (25, 26).
The pro-apoptotic member of proteins contains a BH3 motif, which binds to pro-survival Bcl-2 proteins and prevents them from inhibiting apoptosis. This binding is necessary for the activation of the caspase. In addition, pro-apoptotic Bcl-2 proteins can also bind to autophagy regulators, such as Beclin-1, to induce autophagy, another type of programmed cell death. Viruses have also taken advantage of the Bcl-2 family by encoding their own Bcl-2 proteins, some of which act as apoptosis inhibitors (26).
Fas/FasL: Fas and Fas ligands are crucial in regulating apoptosis. The Fas receptor, also known as CD95 and tumor necrosis factor receptor superfamily member, is located on the surface of various types of cells. It interacts with its corresponding ligand, FasL (also called FasL/CD95L), to initiate a signal transduction pathway that leads to apoptosis. The interaction between FasL and the Fas receptor initiates a series of events that lead to the recruitment and activation of apoptosis-initiating proteases, such as caspase-8 and caspase-10. These proteases subsequently induce apoptosis through various molecules (27).
Inhibitors of Apoptosis Proteins (IAPs): IAPs act as modulators of apoptosis by inhibiting the activity of caspases. This makes them essential in determining cell survival and death, therefore suggesting potential for therapeutic manipulation (25). Overexpression of IAP proteins is often seen in cancers, making them potential targets for therapeutic intervention (28).
Various Types of Programmed Cell Death
Over the last few decades, various regulated cell death pathways have been identified, each relying on a different subset of proteins for their activation and execution. Apoptosis, necroptosis, and pyroptosis are the most-understood forms of cell death. Still, other pathways are being discovered: autophagy-dependent cell death (ADCD), ferroptosis, parthanatos, and, more recently, a Drosophila-specific cell death mechanism called erebosis.
These different forms of cell death are interconnected and share components with other pathways. Studying cell death pathways and their interconnectivity is important for understanding the physiological and mechanistic aspects of these signaling pathways, leading to better disease treatments caused by deregulated or dysfunctional cell death signaling.
Necroptosis: Necroptosis is a type of controlled cell death resembling necrosis, which causes cells to expand and discharge cytoplasmic material (7). The receptor-interacting protein kinases (RIPKs) RIPK1, RIPK3, and Mixed Lineage Kinase Domain-Like Protein (MLKL) are all activated during necroptosis, which is caspase-independent (29).
Various factors, including cancer, viral infections, and tissue damage, can bring it on. It is characterized by cell swelling, membrane rupture, and the release of intracellular contents, including damage-associated molecular patterns (DAMPs) that can draw immune cells.
Necroptosis has been demonstrated to affect cancer prognosis; some researchers suggest that it suppresses tumor growth, while others claim it encourages tumor growth. Additionally, the production of cytokines during necroptosis might cause inflammation and further encourage tumor growth (29).
Pyroptosis: Pyroptosis differs from other cell death types. It is characterized by cell swelling, pore formation on cell membranes, rupture and bubbling of plasma membranes, moderate chromatin condensation, gasdermin (GSDM) cleavage, formation of inflammasomes, and IL-18 and IL-1β release (30).
Intracellular pathogens primarily trigger pyroptosis and facilitate rapid clearance of various infections by removing intracellular replication niches and boosting the host’s defense response (31). It is regulated by inflammasome-associated caspases, such as CASP1, CASP4, CASP5, and CASP11 (mouse), while some caspases associated with apoptosis, such as CASP3 and CASP8 also play a role in pyroptosis (27). It is a crucial mechanism in protecting the host against intracellular pathogens and contributes to antitumor immunity. The induction of pyroptosis can induce potent antitumor activity (29).
Autophagy-dependent cell death (ADCD): Autophagy-dependent cell death (ADCD) is a form of caspase-independent programmed cell death. It’s characterized by an over-activation of autophagy, a cellular degradation process used by eukaryotic cells for maintaining homeostasis and managing lipid metabolism (29).
This over-activation leads to a high degree of self-digestion of cellular material in autolysosomes, moving past the threshold of cell survival24. Autophagy substrates (DAMPs) and pathogen-associated molecular pattern (PAMPs) molecules trigger innate immunity whose elimination through autophagy is necessary for immune homeostasis to protect the cells from exposed membranes and other organelles (29). ADCD occurs during the response of certain cancer cells to specific drugs or natural compounds, showing its relevance in biological and pathological contexts (29, 32).
Ferroptosis: Ferroptosis is regulated cell death dependent on iron and excessive lipid peroxidation, leading to oxidative membrane damage and ruptured plasma membranes. It is distinct from other forms of cell death, regulated by multiple metabolic events, including redox balance, iron handling, and mitochondrial activity (33).
Iron accumulation and lipid peroxidation are the two fundamental processes that trigger ferroptosis (27). In combination with immunotherapy, targeted therapies (inducers or inhibitors) against ferroptosis may exert potent antitumor activity, even in tumors resistant to immune checkpoint inhibitors (29).
Parthanatos: Parthanatos is a form of programmed cell death initiated by PAR polymer, a component of Poly-ADP-ribose polymerase-1 (PARP-1) and is distinct from other cell death processes such as necrosis and apoptosis. It is caused by the accumulation of Poly (ADP ribose) (PAR) and the nuclear translocation of apoptosis-inducing factor (AIF) from mitochondria. Parthanatos’ main mechanism of action is the production of PAR polymer, which leads to NAD+ depletion and the release of apoptosis-inducing factor (AIF), ultimately resulting in cell death.
Other proposed mechanisms of action of parthanatos include mitochondrial permeability transition (PT) and the loss of mitochondrial membrane potential. Understanding parthanatos will help in expanding therapeutic options for PARP-1-related diseases (34).
Erebosis: Erebosis is a newly identified type of cell death discovered in the gut enterocytes of adult Drosophila, or fruit flies. This process differs from known cell death pathways, such as apoptosis, necrosis, or autophagy. It is characterized by the loss of the cytoskeleton, cell adhesion, and organelles. Enterocytes surrounding the erebotic cells often protrude towards them, and old differentiated cells are believed to typically die by apoptosis. Further investigation will be conducted to determine the molecular mechanisms by which erebosis occurs and its role in gut tissue homeostasis (35).
Summary: Exploring the Research on Programmed Cell Death
The exploration of programmed cell death holds profound potential for advancing science and medicine. This fundamental biological event can shed light on diverse domains, including cancer therapy, stem cell research, regenerative medicine, understanding disease mechanisms, drug development, and fundamental biological understanding. Broadly, the course of programmed cell death research promises to enrich Prosci knowledge about the intricacies of cellular life and death, thereby creating opportunities for innovative disease therapies.
Below is a sample of ProSci antibodies related to Apoptosis research. Visit Prosci website to find the right antibodies to help with your research: https://www.prosci-inc.com
| Capases | |
| Cat # | Product |
| 1128 | Caspase-10 Antibody |
| 2071 | Caspase-9 Antibody |
| PM-2015 | Apaf-1 Antibody [2E10] |
| Necroptosis | |
| Cat # | Product |
| 2283 | RIP3 Antibody |
| 3661 | TRAF2 Antibody |
| 3291 | RIPK1 Antibody |
| Ferroptosis | |
| Cat # | Product |
| 14-907 | GPX4 Antibody |
| 13-240 | NRF2 Antibody |
| 16-557 | HSPB1 Antibody, KO Validated |
| Serine Proteases | |
| Cat # | Product |
| 18-708 | GZMB Antibody |
| 22-069 | GZMA Antibody |
| 13-029 | Perforin Antibody |
| Pyroptosis | |
| Cat # | Product |
| 5447 | NALP3 Antibody |
| 16-910 | GSDMD Antibody |
| 3463 | Caspase-1 Antibody |
| 5447 | NALP3 Antibody |
| 16-910 | GSDMD Antibody |
| 3463 | Caspase-1 Antibody |
| Parthantos | |
| Cat # | Product |
| 13-327 | PARP1 Antibody, KO Validated |
| 2267 | AIF Antibody |
| 15-262 | MIF Antibody |
| 13-327 | PARP1 Antibody, KO Validated |
| 2267 | AIF Antibody |
| 15-262 | MIF Antibody |
| BCL-2 | |
| Cat # | Product |
| 3343 | BAD Antibody |
| 3335 | Bcl-2 Antibody |
| 3351 | Bax Antibody |
References:
- Figgett WA, Fairfax K, Vincent FB, Le Page MA, Katik I, Deliyanti D, Quah PS, Verma P, Grumont R, Gerondakis S, Hertzog P, O’Reilly LA, Strasser A, Mackay F. The TACI receptor regulates T-cell-independent marginal zone B cell responses through innate activation-induced cell death. Immunity. 2013 Sep 19;39(3):573-83. doi: 10.1016/j.immuni.2013.05.019. Epub 2013 Sep 5. PMID: 24012421.
- Mahmood SR, Xie X, Hosny El Said N, Venit T, Gunsalus KC, Percipalle P. β-actin dependent chromatin remodeling mediates compartment level changes in 3D genome architecture. Nat Commun. 2021 Sep 2;12(1):5240. doi: 10.1038/s41467-021-25596-2. PMID: 34475390; PMCID: PMC8413440.
- Gottfried Y, Rotem A, Lotan R, Steller H, Larisch S. The mitochondrial ARTS protein promotes apoptosis through targeting XIAP. EMBO J. 2004 Apr 7;23(7):1627-35. doi: 10.1038/sj.emboj.7600155. Epub 2004 Mar 18. PMID: 15029247; PMCID: PMC391065
- Wong RS. Apoptosis in cancer: from pathogenesis to treatment. J Exp Clin Cancer Res. 2011 Sep 26;30(1):87. doi: 10.1186/1756-9966-30-87. PMID: 21943236; PMCID: PMC3197541
- Zhao T, He Q, Xie S, Zhan H, Jiang C, Lin S, Liu F, Wang C, Chen G, Zeng H. A novel Mcl-1 inhibitor synergizes with venetoclax to induce apoptosis in cancer cells. Mol Med. 2023 Jan 19;29(1):10. doi: 10.1186/s10020-022-00565-7. PMID: 36658493; PMCID: PMC9854187.
- Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020 Jul;17(7):395-417. doi: 10.1038/s41571-020-0341-y. Epub 2020 Mar 23. PMID: 32203277; PMCID: PMC8211386.
- Initiation and execution mechanisms of necroptosis: an overview. Cell Death Differ. 2017 Jul;24(7):1184-1195. doi: 10.1038/cdd.2017.65.
- Liu J, Li L. Targeting Autophagy for the Treatment of Alzheimer’s Disease: Challenges and Opportunities. Front Mol Neurosci. 2019 Aug 22;12:203. doi: 10.3389/fnmol.2019.00203. PMID: 31507373; PMCID: PMC6713911.
- Rai SN, Tiwari N, Singh P, Mishra D, Singh AK, Hooshmandi E, Vamanu E, Singh MP. Therapeutic Potential of Vital Transcription Factors in Alzheimer’s and Parkinson’s Disease With Particular Emphasis on Transcription Factor EB Mediated Autophagy. Front Neurosci. 2021 Dec 14;15:777347. doi: 10.3389/fnins.2021.777347. PMID: 34970114; PMCID: PMC8712758.
- Erebosis is a new type of cell death for tissue homeostasis in the Drosophila intestine. PLoS Biol 20(4): e3001614. doi.org/10.1371/journal.pbio.3001614
- The concept of intrinsic versus extrinsic apoptosis. Biochem J. 2022 Feb 11;479(3):357-384. doi: 10.1042/BCJ20210854.
- The proteins and the mechanisms of apoptosis: a mini-review of the fundamentals. Hippokratia. 2007 Jul;11(3):108-13. PMID: 19582203
- Green R, Llambi F. Cell Death Signaling. Cold Spring Harb Perspect Biol. 2015 Dec 1;7(12):a006080. doi: 10.1101/cshperspect.a006080. PMID: 26626938; PMCID: PMC4665079.
- Bots M, Medema JP. Granzymes at a glance. J Cell Sci. 2006 Dec 15;119(Pt 24):5011-4. doi: 10.1242/jcs.03239. PMID: 17158907.
- Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol. 2002 Oct;2(10):735-47. doi: 10.1038/nri911. PMID: 12360212.
- Bots M, Medema JP. Granzymes at a glance. J Cell Sci. 2006 Dec 15;119(Pt 24):5011-4. doi: 10.1242/jcs.03239. PMID: 17158907
- Autophagy, ferroptosis, pyroptosis, and necroptosis in tumor immunotherapy. Signal Transduct Target Ther. 2022 Jun 20;7(1):196. doi: 10.1038/s41392-022-01046-3
- Targeting IAP (inhibitor of apoptosis) proteins for therapeutic intervention in tumors. Curr Cancer Drug Targets. 2008 Mar;8(2):110-7. doi: 10.2174/156800908783769373.
- Caspase activation cascades in apoptosis. Biochem Soc Trans. 2008 Feb;36(Pt 1):1-9. doi: 10.1042/BST0360001.
- Caspase activation pathways: some recent progress. Cell Death Differ. 2009 Jul;16(7):935-8. doi: 10.1038/cdd.2009.59.
- Human caspases: activation, specificity, and regulation. J Biol Chem. 2009 Aug 14;284(33):21777-21781. doi: 10.1074/jbc.R800084200.
- Cell pyroptosis in health and inflammatory diseases. Cell Death Discov. 2022 Apr 11;8(1):191. doi: 10.1038/s41420-022-00998-3.
- Apoptotic and non-apoptotic roles of caspases in neuronal physiology and pathophysiology. Nat Rev Neurosci. 2012 May 18;13(6):395-406. doi: 10.1038/nrn3228.
- Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol. 2002 Oct;2(10):735-47. doi: 10.1038/nri911. PMID: 12360212
- Banjara S, Suraweera CD, Hinds MG, Kvansakul M. The Bcl-2 Family: Ancient Origins, Conserved Structures, and Divergent Mechanisms. Biomolecules. 2020 Jan 12;10(1):128. doi: 10.3390/biom10010128. PMID: 31940915; PMCID: PMC7022251.
- Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol. 2008 Jan;9(1):47-59. doi: 10.1038/nrm2308. PMID: 18097445
- Yu WR, Fehlings MG. Fas/FasL-mediated apoptosis and inflammation are key features of acute human spinal cord injury: implications for translational, clinical application. Acta
- Targeting IAP (inhibitor of apoptosis) proteins for therapeutic intervention in tumors. Curr Cancer Drug Targets. 2008 Mar;8(2):110-7. doi: 10.2174/156800908783769373.
- Research progresses of molecular mechanism of pyroptosis and its related diseases. Immunobiology. 2020 Mar;225(2):151884. doi: 10.1016/j.imbio.2019.11.019.
- Cell pyroptosis in health and inflammatory diseases. Cell Death Discov. 2022 Apr 11;8(1):191. doi: 10.1038/s41420-022-00998-3.
- Research progresses of molecular mechanism of pyroptosis and its related diseases. Immunobiology. 2020 Mar;225(2):151884. doi: 10.1016/j.imbio.2019.11.019.
- Organelle-specific mechanisms of drug-induced autophagy-dependent cell death. Matrix Biol. 2021 Jun;100-101:54-64. doi: 10.1016/j.matbio.2020.12.003
- Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021 Apr;22(4):266-282. doi: 10.1038/s41580-020-00324-8
- David KK, Andrabi SA, Dawson TM, Dawson VL. Parthanatos, a messenger of death. Front Biosci (Landmark Ed). 2009 Jan 1;14(3):1116-28. doi: 10.2741/3297. PMID: 19273119; PMCID: PMC4450718.
- Erebosis is a new type of cell death for tissue homeostasis in the Drosophila intestine. PLoS Biol 20(4): e3001614. doi.org/10.1371/journal.pbio.3001614
Molecular Mechanisms Regulating Necroptosis
Insights into the Molecular Mechanisms Regulating Necroptosis
Introduction
Necroptosis, a form of regulated cell death, has gained significant attention recently due to its involvement in various physiological and pathological processes. Necroptosis is a specialized form of programmed cell death that is tightly controlled by various signaling pathways, mediators, and immune responses. Interferon-γ (IFN-γ) and Toll-like receptor 4 (TLR4) signaling pathways have been identified as critical regulators of necroptosis in macrophages. Understanding the underlying molecular mechanisms that govern necroptosis is essential for unraveling its role in immune responses and disease pathogenesis.
This white paper introduces the recent findings in molecular mechanisms that govern necroptosis as they relate to receptor-interacting protein kinases (RIPK) and provides examples of ProSci’s rabbit anti-RIPK3 antibody (ProSci Cat #: 2283) when used to analyze the expression of RIPK3 in various cell lines by Western Blot.
Genesee Scientific’s ProSci portfolio brings more than 25 years of antibody expertise in developing high quality single domain Variable Heavy domain of Heavy chain (VHH), recombinant, monoclonal, and polyclonal antibodies. In addition, Prosci custom antibody services provide the flexibility to tailor antibody development and production for almost any need.
“RIPK3 and MLKL: Essential Players in IFN-γ Induced PS Exposure and Necroptosis”
The paper published in 2019 by Chen et al. titled “Interferon induces the cell surface exposure of phosphatidylserine (PS) by activating the protein MLKL in the absence of caspase-8 activity” presents research findings on the role of RIPK3 in IFN-γ-induced PS exposure in mouse embryonic fibroblasts (MEFs). The authors demonstrated that RIPK3 is necessary for PS exposure in MEFs. At the same time, RIPK3-activated oligomerization of the mixed lineage kinase domain-like protein (MLKL) protein is essential for IFN-γ induced PS exposure and the execution of necroptotic cell death in MEFs (1).
Necroptosis is a non-apoptotic, regulated form of necrosis triggered by inducers such as tumor necrosis factor (TNF) binding to their receptors (2). It requires the activity of RIPK1 and RIPK3 and is mediated by activating MLKL. (3) Interferon (IFN) is classified into two groups, type I and type II, and both types of IFN induce the expression of hundreds of specific genes (4,5).
This paper demonstrated that RIPK3 is necessary for IFN-γ-induced PS exposure and necroptosis in RIPK3-reconstituted iC8KO MEFs, immortalized caspase-8 knockout (iC8KO) mouse-derived embryonic fibroblasts with exogenously expressed RIPK3, and that MLKL is phosphorylated and oligomerized in IFN-γ treated MEFs before necroptosis. Furthermore, IFN-γ-induced PS exposure and necroptosis depend on RIPK1, RIPK3, and MLKL in various cells.
This research highlights the role of RIPK3, MLKL, and IFN-γ in inducing PS exposure and necroptosis in MEFs and provides insights into the mechanisms underlying IFN-γ-induced PS exposure and the potential functions of MLKL in this process. Aided by the use of ProSci’s rabbit anti-RIPK3 antibody, these findings could lead to the development of new therapeutic strategies for targeting necroptosis in diseases such as cancer and infection.
“Impact of RIPK1 K45A Mutation on Necroptosis, Cytokine Signaling, and Immune Response: An Altered Landscape of Inflammatory and Infection Susceptibility”
The paper published in 2016 by Shutinoski et al. titled “K45A mutation of RIPK1 results in poor necroptosis and cytokine signaling in macrophages, which impacts inflammatory responses in vivo” investigated the role of the kinase domain of RIPK1 in necroptosis and cytokine signaling in macrophages, and its consequent impact on inflammatory response in vivo. It also evaluated the effect of a K45A mutation of RIPK1 on the necroptosis of macrophages and the activation of inflammatory response (7).
RIPK1 is essential for the induction of necroptosis and is triggered by the engagement of various Toll-like receptors (TLRs) and cytokine receptors (8,9). The phosphorylation of RIPK1 leads to the interaction of RIPK1 and RIPK3, which is necessary for necroptosis to occur (10). RIPK3 can also induce necroptosis independently of RIPK1 in certain cases (11). Furthermore, the MLKL protein is also essential for necroptosis, as its phosphorylation results in its oligomerization and translocation to the cell membrane, disrupting cellular integrity (12). Thus, RIPK1 and MLKL proteins play vital roles in the induction of necroptosis through their respective interactions and phosphorylation.
In their study, the authors employed the RIPK1^K45A strain of mice to elucidate the crucial role of RIPK1 kinase activity in facilitating macrophage necroptosis. Through their investigations, they observed that the RIPK1^K45A mutation led to a decline in cytokine production, a reduction in the phosphorylation of RIPK1 and MLKL proteins, and an elevation in cell survival after treatments involving the combination of lipopolysaccharide (LPS) and a cell-permeable pan-caspase inhibitor (zVAD) as well as tumor necrosis factor alpha (TNFα) and zVAD. LPS/zVAD treatment was employed to induce necroptosis in macrophages, where macrophages were exposed to LPS along with zVAD. Similarly, TNFα/zVAD treatment entailed the application of TNFα, an inflammatory cytokine, in conjunction with zVAD to induce necroptosis in macrophages.
Additionally, the researchers uncovered the regulatory influence of RIPK1 on TNFα signaling and its involvement in the transcriptional regulation of macrophage inflammatory protein-1 alpha (MIP-1α) and interleukin-1 alpha (IL-1α). Furthermore, the researchers made an intriguing observation that caspase-8 inhibitors heightened the occurrence of necroptosis in macrophages, particularly when the p38MAPK pathway was suppressed.
This study demonstrated that lysine-45 of RIPK1 is required for RIPK1 auto-phosphorylation and cell death of macrophages induced by various stimuli, including LPS and TNF-α. RIPK1K45A macrophages were resistant to cell death induced by these stimuli and showed reduced phosphorylation of STAT1 in response to LPS/ZVAD and TNF-α/ZVAD treatments. These results suggest that RIPK1K45A is involved in regulating cell death downstream of TNF-R engagement and plays a role in preventing excessive cell death in response to various stimuli.
The study showed that phosphorylation of RIPK3 was attenuated in RIPK1K45A macrophages, indicating this kinase domain’s important role in promoting macrophage necroptosis. The RIPK3 antibody was used to identify and measure the levels of RIPK3, as well as its interaction with RIPK1 and MLKL using western blot. Overall, the results suggest that the RIPK3 antibody was essential in characterizing RIPK1 kinase activity and its role in regulating necroptosis, inflammatory response, and cytokine signaling in macrophages.
Conclusion
These investigations provide significant insights into the roles of RIPK3, MLKL, and IFN-γ in the initiation of phosphatidylserine presentation and necroptosis in mouse embryonic fibroblasts (MEFs). Furthermore, it underscores the crucial roles of RIPK3 and MLKL in the IFN-γ-mediated phosphatidylserine display and the necroptosis pathway.
In addition, a mutation in RIPK1, specifically RIPK1K45A, hinders the necroptosis of macrophages and the ignition of the inflammatory response. This research emphasizes the role of the RIPK1 kinase domain in initiating necroptosis and cytokine signaling within macrophages and the subsequent effects on in vivo inflammatory responses.
The ProSci rabbit anti-RIPK3 antibody was employed in this investigation to examine the expression of RIPK3 across different cell lines, playing a pivotal role in delineating RIPK1 kinase activity and its regulatory functions in necroptosis, inflammatory reactions, and cytokine signaling in macrophages. The results of these investigations have the potential to facilitate additional studies into the functions of RIPK3 and MLKL in the process of phosphatidylserine exposure and necroptosis, thereby advancing Prosci comprehension of the underlying mechanisms of cell death in the immune system that leads to the development of new therapeutic strategies for targeting necroptosis in diseases such as cancer and infection.
Visit Prosci website to learn more about anti-RIPK3 antibody and related antibodies needed to help with your research: https://www.prosci-inc.com
References:
- Chen J, Kuroki S, Someda M, Yonehara S. Interferon-γ induces the cell surface exposure of phosphatidylserine by activating the protein MLKL in the absence of caspase-8 activity. J Biol Chem. 2019 Aug 9;294(32):11994-12006. doi: 10.1074/jbc.RA118.007161. Epub 2019 Jun 19. PMID: 31217278; PMCID: PMC6690710.
- Dondelinger Y, Hulpiau P, Saeys Y, Bertrand MJM, Vandenabeele P. An evolutionary perspective on the necroptotic pathway. Trends Cell Biol. 2016 Oct;26(10):721-732. doi: 10.1016/j.tcb.2016.06.004. Epub 2016 Jun 28. PMID: 27368376.
- Cho YS, Challa S, Moquin D, Genga R, Ray TD, Guildford M, Chan FK. Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell. 2009 Jun 12;137(6):1112-23. doi: 10.1016/j.cell.2009.05.037. PMID: 19524513; PMCID: PMC2727676.
- Pestka S, Krause CD, Walter MR. Interferons, interferon-like cytokines, and their receptors. Immunol Rev. 2004 Dec;202:8-32. doi: 10.1111/j.0105-2896.2004.00204.x. PMID: 15546383.
- Borden EC, Sen GC, Uze G, Silverman RH, Ransohoff RM, Foster GR, Stark GR. Interferons at age 50: past, current and future impact on biomedicine. Nat Rev Drug Discov. 2007 Dec;6(12):975-90. doi: 10.1038/nrd2422. PMID: 18049472; PMCID: PMC7097588.
- Zargarian S, Shlomovitz I, Erlich Z, Hourizadeh A, Ofir-Birin Y, Croker BA, Regev-Rudzki N, Edry-Botzer L, Gerlic M. Phosphatidylserine externalization, “necroptotic bodies” release, and phagocytosis during necroptosis. PLoS Biol. 2017 Jun 26;15(6):e2002711. doi: 10.1371/journal.pbio.2002711. PMID: 28650960; PMCID: PMC5501695.
- Shutinoski B, Alturki NA, Rijal D, Bertin J, Gough PJ, Schlossmacher MG, Sad S. K45A mutation of RIPK1 results in poor necroptosis and cytokine signaling in macrophages, which impacts inflammatory responses in vivo. Cell Death Differ. 2016 Oct;23(10):1628-37. doi: 10.1038/cdd.2016.51. Epub 2016 Jun 3. PMID: 27258786; PMCID: PMC5041191.
- Kelliher MA, Grimm S, Ishida Y, Kuo F, Stanger BZ, Leder P. The death domain kinase RIP mediates the TNF-induced NF-kappaB signal. Immunity. 1998 Mar;8(3):297-303. doi: 10.1016/s1074-7613(00)80535-x. PMID: 9529147.
- Vandenabeele P, Galluzzi L, Vanden Berghe T, Kroemer G. Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat Rev Mol Cell Biol. 2010 Oct;11(10):700-14. doi: 10.1038/nrm2970. Epub 2010 Sep 8. PMID: 20823910.
- Cho YS, Challa S, Moquin D, Genga R, Ray TD, Guildford M, Chan FK. Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell. 2009 Jun 12;137(6):1112-23. doi: 10.1016/j.cell.2009.05.037. PMID: 19524513; PMCID: PMC2727676.
- Upton JW, Kaiser WJ, Mocarski ES. Virus inhibition of RIP3-dependent necrosis. Cell Host Microbe. 2010 Apr 22;7(4):302-313. doi: 10.1016/j.chom.2010.03.006. PMID: 20413098; PMCID: PMC4279434.
- Sun L, Wang H, Wang Z, He S, Chen S, Liao D, Wang L, Yan J, Liu W, Lei X, Wang X. Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase. Cell. 2012 Jan 20;148(1-2):213-27. doi: 10.1016/j.cell.2011.11.031. PMID: 22265413.
PD-1: Expanding Beyond Cancer Research
PD-1: Expanding Beyond Cancer Research Introduction to PD-1
Introduction to PD-1
There has been significant interest in the role of programmed cell death protein 1 (PD-1) and its ligand programmed death-ligand 1 (PD-L1) in modulating immune responses. The emergence of PD-1-targeted therapies has revolutionized cancer treatment, leading to a better understanding of immune checkpoint mechanisms and their uses in various research fields.
This paper offers a review of significant publications on the use of PD-1 antibodies in cancer, immunology, and Alzheimer’s disease research. We will highlight the importance of PD-1 antibodies in resolving the complicated interactions between the immune system and pathological conditions as well as their potential to create novel therapeutic approaches.
Download the white paper here.
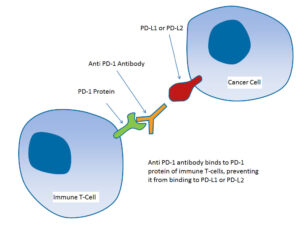
Enhancing Immunology Research & Therapy Development
The ProSci PD-1 antibody has been used in a variety of cancer, immunology, and Alzheimer’s disease investigations. Researchers have successfully detected PD-1 expression in a several of study settings with the highly specific rabbit polyclonal anti-PD-1 antibody (ProSci Cat #: 4065, Genesee Cat #: GS1-681).
The ProSci PD-1 antibody has been found to be effective in improving our understanding of the immune system’s participation in many pathological conditions, and it has aided in the development of novel therapeutic strategies.
Learn More: https://www.prosci-inc.com/product/pd-1-antibody-4065/
Figure 1: ProSci PD-1 Antibody (ProSci Cat #: 4065, Genesee Cat #: GS1-681) Western Blot Validation in Human and Mouse Cell Lines
Loading: 15μg of lysates per lane. Antibodies: PD-1 (ProSci Cat #: 4065, Genesee Cat #: GS1-681) (4μg/mL), 1h incubation at RT in 5% NFDM/TBST
Secondary: Goat anti-rabbit IgG HRP conjugate at 1:10000 dilution
Figure 2: ProSci PD-1 Antibody (ProSci Cat #: 4065, Genesee Cat #: GS1-681) Immunohistochemistry Validation of PD-1in Human Tonsil Tissue Tissue was fixed with formaldehyde and blocked with 10% serum for 1h at RT; antigen retrieval was by heat mediation with a citrate buffer (pH6). Samples were incubated with primary antibody overnight at 4°C. A goat anti-rabbit IgG H&L (HRP) at 1:250 was used as a secondary antibody and was counter-stained with Hematoxylin.
Utilization of ProSci PD-1 Antibody in Immunology Research:
The ProSci PD-1 antibody facilitated key insights into how BCG vaccination and GSH enhancement modulate PD-1 expression, bolstering its versatility.
In the study “Elucidating the Efficacy of the Bacille Calmette-Guerin Vaccination in Conjunction with
First Line Antibiotics and Liposomal Glutathione” 1, researchers investigated the combined effects of
BCG (Bacille Calmette-Guerin) vaccination and ex-vivo glutathione (GSH) enhancement on improving
immune responses against Mycobacterium tuberculosis (M. tb) infection.
They examined immune response effectiveness, CD4+ and CD8+ T cells in granulomas, liposomal glutathione (L-GSH) response, antibiotic levels, PD-1 expression, and autophagy induction. The results demonstrated that BCG vaccination, along with GSH enhancement, prevented CD4+ and CD8+ T cell loss in granulomas, improved M. tb infection control, reduced PD-1 expression, and increased autophagy and cytokine production (IFN-γ and TNF-α).
The ProSci rabbit polyclonal anti-PD-1 antibody (ProSci Cat #: 4065, Genesee Cat #: GS1-681) was used to measure PD-1 expression in FFPE tissue samples. The results revealed a decrease in PD-1 expression due to BCG vaccination in immune responses against M. tb infection.
Utilization of ProSci PD-1 and PD-L1 Antibodies in Alzheimer’s Disease Research:
ProSci PD-1 and PD-L1 antibodies uniquely illuminated the PD-1/PD-L1
axis’ role in Alzheimer’s pathology and neuroinflammation.
In the study “Microglial PD-1 stimulation by astrocytic PD-L1 suppresses neuroinflammation and
Alzheimer’s disease pathology” 2, researchers investigated the role of the PD-1/PD-L1 immune
checkpoint in Alzheimer’s disease (AD). They demonstrated upregulated expression of PD-L1 in
astrocytes and PD-1 in microglia surrounding amyloid plaques in AD patients and the APP/PS1 mouse
model.
Deletion of microglial PD-1 resulted in increased amyloid-beta (Aβ) deposition, reduced microglial Aβ uptake, and decreased expression of the Aβ receptor CD36 on microglia. These findings suggest that ineffective immune regulation by the PD-1/PD-L1 axis contributes to Aβ plaque deposition during chronic neuroinflammation in AD.
The ProSci rabbit polyclonal anti-PD-1 (ProSci Cat #: 4065, Genesee Cat #: GS1-681) and anti-PD-L1
(ProSci Cat #: 4059, Genesee Cat #: GS1-679) antibodies were used to measure the expression levels of
PD-1 and PD-L1 in FFPE tissue samples from Alzheimer’s patients.
Utilization of ProSci PD-1 Antibody in Cancer Research:
ProSci’s PD-1 antibody facilitated a detailed analysis of rare D-LPHL, underscoring diagnostic versatility.
In the study “Diffuse variant of lymphocyte-predominant Hodgkin lymphoma: a diagnostic challenge” 3, researchers presented a rare case of diffuse variant nodular lymphocyte-predominant Hodgkin lymphoma (D-LPHL) in a 35-year-old Caucasian female with left cervical and right axillary lymphadenopathy. Histopathological examination revealed large atypical lymphoid cells with a characteristic phenotype. Immunohistochemical studies confirmed the diffuse nature of the lymphoid proliferation, and the associated T cell microenvironment showed increased CD57(+) T cells. The report discusses the diagnostic criteria and provides a review of the literature on the differential diagnosis of this rare condition.
The ProSci rabbit polyclonal anti-PD-1 (ProSci Cat #: 4065, Genesee Cat #: GS1-681) antibody was used to detect the expression of PD-1 in FFPE tissue sample from this rare case of diffuse variant nodular lymphocyte-predominant Hodgkin lymphoma. The study highlighted the presence of PD-1-positive T cells exhibiting rosette formation around LP-type HL cells.
Research Tools
This paper highlights the significance of the ProSci PD-1 antibody in cancer, immunology, and Alzheimer’s disease research, demonstrating its effectiveness in understanding the immune system’s
role in various pathological conditions. The paper showcases the antibody’s potential in developing
novel therapeutic approaches and resolving complex interactions between the immune system and
diseases by analyzing key publications.
For more information on PD-1 research tools and other high performance antibody tools, contact ProSci Antibodies at customercare@prosci-inc.com or visit www.geneseesci.com. or customerservice@biolinkk.com
Learn more about PD-1 Antibody: https://www.prosci-inc.com/product/pd-1-antibody-4065/
OVERVIEW OF BISULFITE CONVERSION
Learn More About Bisulfite Conversion
OVERVIEW OF BISULFITE CONVERSION
When starting with bisulfite conversion, there are a number of considerations to keep in mind to ensure optimal performance and subsequent accuracy of the DNA methylation analysis. Treatment with bisulfite is inherently a harsh process that dramatically changes both the chemical makeup and physical properties of the DNA. Input DNA transforms from a large, stable, double-stranded molecule to a collection of randomly fragmented, single-stranded fragments having almost all cytosines completely changed to uracil.
These changes dramatically influence quantification, quality assessment, amplification, and analysis of the bisulfite-converted DNA. These points and the special considerations they require with respect to UV spectrophotometry (NanoDrop), agarose gel electrophoresis, and PCR are highlighted below.
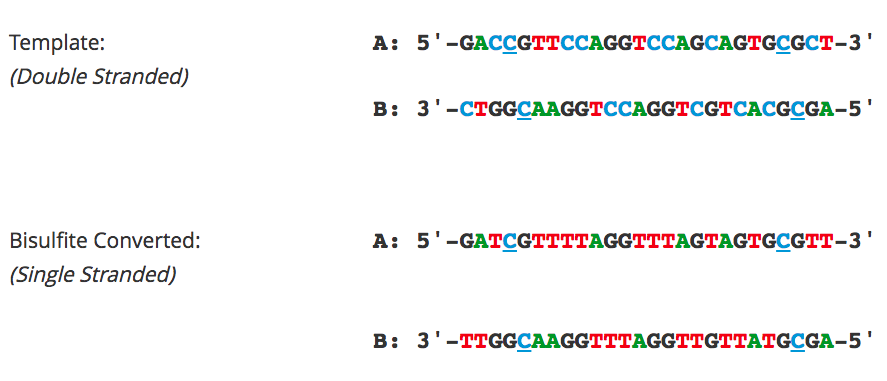
QUANTIFICATION OF BISULFITE-CONVERTED DNA

Converted DNA should be quantitated as RNA using a UV spectrophotometer (NanoDrop) with Ab260 nm 1.0 = 40 µg/ml. When determining the recovery of the bisulfite-converted DNA, two primary factors need to be considered 1.) The intactness of the starting material, and 2.) RNA contamination. The quality of the DNA, in terms of size, used as input for bisulfite conversion is the most important factor when assessing recovery. Degraded starting material will lead to increased sample loss during the bisulfite conversion process. Additionally, RNA contamination will contribute significantly to the Ab260 nm resulting in an overestimation of DNA quantity. RNA carried over into the conversion is lost making the yields appear low compared to the input. Importantly, the conversion is not compromised in either of the above cases.
ASSESSING QUALITY OF BISULFITE-CONVERTED DNA
Agarose gel electrophoresis (2% gel with 100 bp marker) of converted DNA can be used to gauge recovery and fragmentation. Most often nothing will be visible in the gel when it is first removed from the gel tank, this is normal and results from the DNA being almost entirely single stranded following the conversion. Chilling the gel for several minutes in an ice bath will force enough base-pairing to allow intercalation of the ethidium bromide for the DNA to be visible. The converted DNA will run as a smear, generally from > 1,500 down to 100 bp. It is important that enough material be loaded into the gel to allow visualization: typically about 100 ng/well is sufficient.

The Challenges of Bisulfite-Converted DNA – DNA fragmentation and limited base pairing makes it difficult to see the DNA following bisulfite conversion. Samples of bisulfite converted human genomic DNA were run in a 2% agarose gel with a 100 bp marker visualized normally (left panel) and after chilling in an ice bath (right panel).
BISULFITE PCR AND PRIMER DESIGN
Bisulfite PCR is the most common technique used for methylation analysis of bisulfite-converted DNA and also the most susceptible to mishap. Keep in mind the DNA will be significantly fragmented, as the strands are no longer complementary, and almost completely devoid of cytosine. Primer design is the key to successful bisulfite PCR. Unlike normal PCR, bisulfite PCR primers need to be long (usually between 26-30 bases) and the amplicon size should be relatively short (between 150-300 bp). Ideally, the primers should not contain CpG sites, however, if they are necessary locate them at the 5′-end of the primer with a mixed base at the cytosine position. It is also important to note that only one strand of the bisulfite-converted template will be amplified by any given primer set. Only the reverse primer will actually bind to the target DNA, which in turn will generate template for the forward primer to anneal. Usually, 35 to 40 cycles are required for successful amplification. Hot start polymerases are strongly recommended as non-specific amplification is relatively common with bisulfite-converted DNA due to it being AT-rich. Annealing temperatures between 55-60°C typically work well, and an annealing temperature gradient should be run with every new primer set to ensure optimal amplification of the specific target.
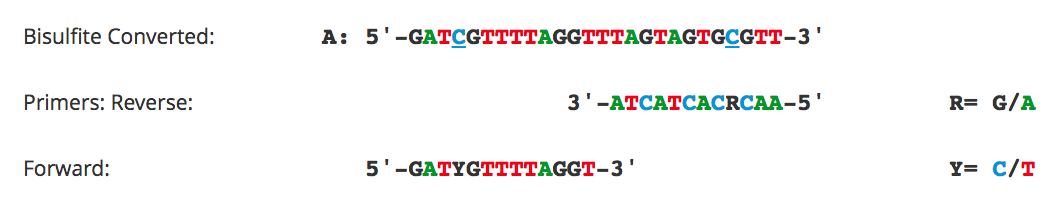
The Challenges of Bisulfite-Converted DNA – DNA fragmentation and limited base pairing makes it difficult to see the DNA following bisulfite conversion. Samples of bisulfite converted human genomic DNA were run in a 2% agarose gel with a 100 bp marker visualized normally (left panel) and after chilling in an ice bath (right panel).
PRIMER DESIGN FOR METHYLATION SPECIFIC PCR (MSP)
Methylation Specific PCR (MSP) relies on amplification to assess the methylation status at specific CpG sites. Success with this system depends on the differential amplification of the template using methylated (M) and non-methylated (U) primer sets. While most of the considerations for primer design are identical to those for bisulfite PCR, treatment of CpG sites within the primer is completely different. For MSP it is necessary to locate CpG sites at the 3′-end of the primers with cytosines in the methylated (M) primers and thymines in the non-methylated (U) primers.
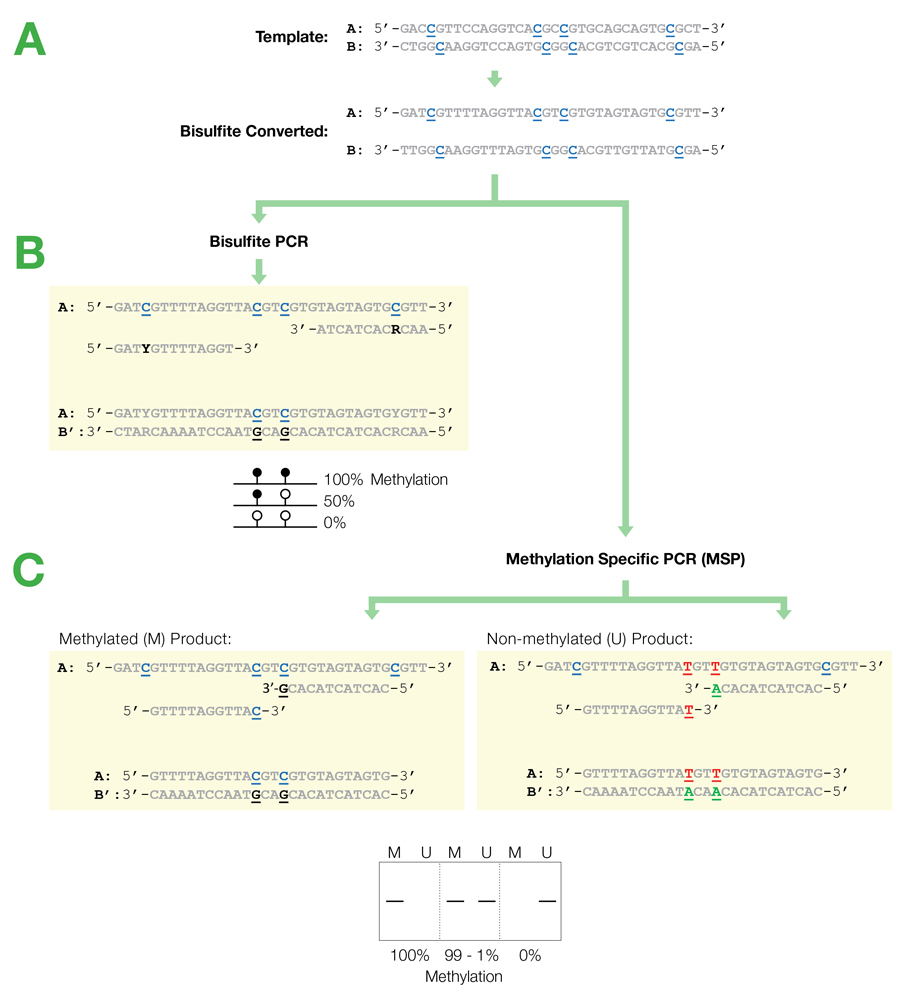
Flowchart of primer design for bisulfite PCR and Methylation Specific PCR (MSP). (A) Following bisulfite treatment, the two converted strands of the DNA template are no longer complementary. It is important to note that only one strand will be amplified with any given primer set. This is illustrated by the following examples: (B) Primers for Bisulfite PCR are designed for subsequent sequencing and analysis of cytosines within the amplicon. CpG sites within the primers should be avoided or located at the 5′-end with a mixed base at the cytosine position (Y= C/T, R= G/A). Sequencing data is commonly represented by a “lollipop” plot where closed circles represent methylated cytosine positions and open circles non-methylated ones. (C) Primers for Methylation Specific PCR (MSP) are designed to target and assess the methylation status at specific CpG sites. CpG sites within the primers must be located at the 3’-end to increase their specificity to methylated (M) or non-methylated (U) templates. Completely methylated or non-methylated templates will generate a single amplicon from only their representative primer set following MSP. Samples with mixed methythation, will be amplified by both primer sets.
BISULFITE SEQUENCING
Bisulfite sequencing is still among the most common techniques used for analyzing bisulfite-converted DNA and provides single base resolution across the entire amplicon. Cloning followed by sequencing with vector-specific primers is recommended to obtain the best sequencing results for quantification of methylation. Direct sequencing of bisulfite PCR products is not recommended as it generally yields poor sequencing quality and will confound quantification of partially methylated sites. Pyrosequencing can be used as an alternative to direct sequencing.
NEXT-GEN BISULFITE SEQUENCING
Next-Gen sequencing-based applications such as Reduced Representation Bisulfite Sequencing (RRBS) and Whole-Genome Bisulfite Sequencing (WGBS) are increasingly utilized to obtain genome-wide single base resolution methylation analysis and to identify putative biomarker candidates. In both workflows, conversion efficiency is of paramount concern, as even relatively small variations in efficiency can effect thousands of individual sites. If library preparation is to be performed prior to bisulfite conversion, it is important that the adaptors added to the fragmented DNA be methylated to preserve their sequence. Use of proper control DNAs are also recommended if non-CpG methylation is a factor in the samples (e.g., in plants) and to account for the inability of bisulfite conversion to differentiate between methylation (5-mC) and hydroxymethylation (5-hmC).
Methylation Plot from Reduced Representation Bisulfite Sequencing (RRBS). Methylation Plot from Reduced Representation Bisulfite Sequencing (RRBS). Data shows the relative percentage of methylation at individual CpG sites in mouse DNA. Methylation percentage is shown across a ~3 Mb region of mouse chromosome 19. Bisulfite sequencing libraries were prepared using mouse genomic DNA prepped with the Genomic Clean & Concentrator (D4010, D4011 – Zymo Research) and bisulfite converted using EZ DNA Methylation technology prior to Next-Gen sequencing.